The urea cycle (also known as the ornithine cycle) is a cycle of biochemical reactions that produces urea (NH2)2CO from ammonia (NH3). Animals that use this cycle, mainly amphibians and mammals, are called ureotelic.
The urea cycle converts highly toxic ammonia to urea for excretion.[1] This cycle was the first metabolic cycle to be discovered (Hans Krebs and Kurt Henseleit, 1932), five years before the discovery of the TCA cycle. This cycle was described in more detail later on by Ratner and Cohen. The urea cycle takes place primarily in the liver and, to a lesser extent, in the [[kidney].
Function
Amino acid catabolism results in waste ammonia. All animals need a way to excrete this product. Most aquatic organisms, or ammonotelic organisms, excrete ammonia without converting it.[1] Organisms that cannot easily and safely remove nitrogen as ammonia convert it to a less toxic substance, such as urea, via the urea cycle, which occurs mainly in the liver. Urea produced by the liver is then released into the bloodstream, where it travels to the kidneys and is ultimately excreted in urine. The urea cycle is essential to these organisms, because if the nitrogen or ammonia is not eliminated from the organism it can be very detrimental.[2] In species including birds and most insects, the ammonia is converted into uric acid or its urate salt, which is excreted in solid form. Further, the urea cycle consumes acidic waste carbon dioxide by combining it with the basic ammonia, helping to maintain a neutral pH.
Reactions
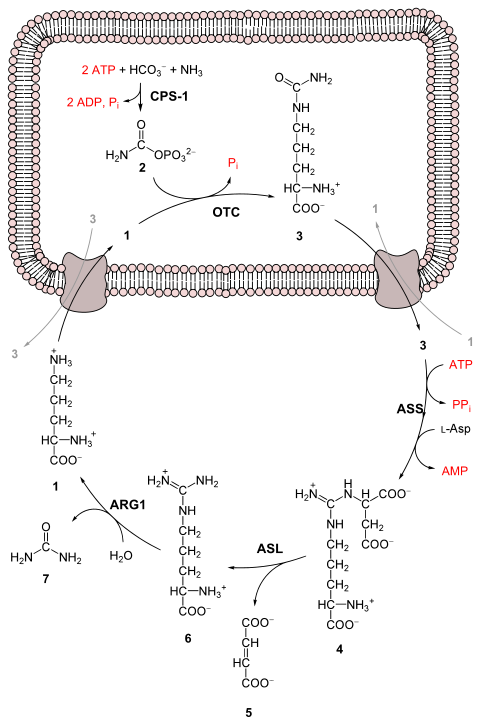
The entire process converts two amino groups, one from NH+ 4 and one from aspartate, and a carbon atom from HCO− 3, to the relatively nontoxic excretion product urea.[3] This occurs at the cost of four "high-energy" phosphate bonds (3 ATP hydrolyzed to 2 ADP and one AMP). The conversion from ammonia to urea happens in five main steps. The first is needed for ammonia to enter the cycle and the following four are all a part of the cycle itself. To enter the cycle, ammonia is converted to carbamoyl phosphate. The urea cycle consists of four enzymatic reactions: one mitochondrial and three cytosolic.[1][4] This uses 6 enzymes.[3][4][5]
Before the urea cycle begins ammonia is converted to carbamoyl phosphate. The reaction is catalyzed by carbamoyl phosphate synthetase I and requires the use of two ATP molecules.[1] The carbamoyl phosphate then enters the urea cycle.
Steps of the urea cycle
Carbamoyl phosphate is converted to citrulline. With catalysis by ornithine transcarbamylase, the carbamoyl phosphate group is donated to ornithine and releases a phosphate group.[1]
Arginine is cleaved by arginase to form urea and ornithine. The ornithine is then transported back to the mitochondria to begin the urea cycle again.[1][4]
Overall reaction equation
In the first reaction, NH+ 4 + HCO− 3 is equivalent to NH3 + CO2 + H2O.
Since fumarate is obtained by removing NH3 from aspartate (by means of reactions 3 and 4), and PPi + H2O → 2 Pi, the equation can be simplified as follows:
Note that reactions related to the urea cycle also cause the production of 2 NADH, so the overall reaction releases slightly more energy than it consumes. The NADH is produced in two ways:
One NADH molecule is produced by the enzyme glutamate dehydrogenase in the conversion of glutamate to ammonium and α-ketoglutarate. Glutamate is the non-toxic carrier of amine groups. This provides the ammonium ion used in the initial synthesis of carbamoyl phosphate.
The fumarate released in the cytosol is hydrated to malate by cytosolic fumarase. This malate is then oxidized to oxaloacetate by cytosolic malate dehydrogenase, generating a reduced NADH in the cytosol. Oxaloacetate is one of the keto acids preferred by transaminases, and so will be recycled to aspartate, maintaining the flow of nitrogen into the urea cycle.
The two NADH produced can provide energy for the formation of 5 ATP (cytosolic NADH provides 2.5 ATP with the malate-aspartate shuttle in human liver cell), a net production of two high-energy phosphate bond for the urea cycle. However, if gluconeogenesis is underway in the cytosol, the latter reducing equivalent is used to drive the reversal of the GAPDH step instead of generating ATP.
The fate of oxaloacetate is either to produce aspartate via transamination or to be converted to phosphoenolpyruvate, which is a substrate for gluconeogenesis.
Products of the urea cycle
As stated above many vertebrates use the urea cycle to create urea out of ammonium so that the ammonium does not damage the body. Though this is helpful, there are other effects of the urea cycle. For example: consumption of two ATP, production of urea, generation of H+, the combining of HCO−3 and NH+4 to forms where it can be regenerated, and finally the consumption of NH+4.[6]
Regulation
N-Acetylglutamic acid
The synthesis of carbamoyl phosphate and the urea cycle are dependent on the presence of N-acetylglutamic acid (NAcGlu), which allosterically activates CPS1. NAcGlu is an obligate activator of carbamoyl phosphate synthetase.[7] Synthesis of NAcGlu by N-acetylglutamate synthase (NAGS) is stimulated by both Arg, allosteric stimulator of NAGS, and Glu, a product in the transamination reactions and one of NAGS's substrates, both of which are elevated when free amino acids are elevated. So Glu not only is a substrate for NAGS but also serves as an activator for the urea cycle.
Substrate concentrations
The remaining enzymes of the cycle are controlled by the concentrations of their substrates. Thus, inherited deficiencies in cycle enzymes other than ARG1 do not result in significant decreases in urea production (if any cycle enzyme is entirely missing, death occurs shortly after birth). Rather, the deficient enzyme's substrate builds up, increasing the rate of the deficient reaction to normal.
The anomalous substrate buildup is not without cost, however. The substrate concentrations become elevated all the way back up the cycle to NH+ 4, resulting in hyperammonemia (elevated [NH+ 4]P).
Although the root cause of NH+ 4 toxicity is not completely understood, a high [NH+ 4] puts an enormous strain on the NH+ 4-clearing system, especially in the brain (symptoms of urea cycle enzyme deficiencies include intellectual disability and lethargy). This clearing system involves GLUD1 and GLUL, which decrease the 2-oxoglutarate (2OG) and Glu pools. The brain is most sensitive to the depletion of these pools. Depletion of 2OG decreases the rate of TCAC, whereas Glu is both a neurotransmitter and a precursor to GABA, another neurotransmitter. (p.734)
Link with the citric acid cycle
The urea cycle and the citric acid cycle are independent cycles but are linked. One of the nitrogen atoms in the urea cycle is obtained from the transamination of oxaloacetate to aspartate.[8] The fumarate that is produced in step three is also an intermediate in the citric acid cycle and is returned to that cycle.[8]
Urea cycle disorders
Urea cycle disorders are rare and affect about one in 35,000 people in the United States.[9]Genetic defects in the enzymes involved in the cycle can occur, which usually manifest within a few days after birth.[2] The recently born child will typically experience varying bouts of vomiting and periods of lethargy.[2] Ultimately, the infant may go into a coma and develop brain damage.[2] New-borns with UCD are at a much higher risk of complications or death due to untimely screening tests and misdiagnosed cases. The most common misdiagnosis is neonatal sepsis. Signs of UCD can be present within the first 2 to 3 days of life, but the present method to get confirmation by test results can take too long.[10] This can potentially cause complications such as coma or death.[10]
Urea cycle disorders may also be diagnosed in adults, and symptoms may include delirium episodes, lethargy, and symptoms similar to that of a stroke.[11] On top of these symptoms, if the urea cycle begins to malfunction in the liver, the patient may develop cirrhosis.[12] This can also lead to sarcopenia (the loss of muscle mass).[12] Mutations lead to deficiencies of the various enzymes and transporters involved in the urea cycle, and cause urea cycle disorders.[1] If individuals with a defect in any of the six enzymes used in the cycle ingest amino acids beyond what is necessary for the minimum daily requirements, then the ammonia that is produced will not be able to be converted to urea. These individuals can experience hyperammonemia, or the build-up of a cycle intermediate.
All urea cycle defects, except OTC deficiency, are inherited in an autosomal recessive manner. OTC deficiency is inherited as an X-linked recessive disorder, although some females can show symptoms. Most urea cycle disorders are associated with hyperammonemia, however argininemia and some forms of argininosuccinic aciduria do not present with elevated ammonia.
Additional images
Urea cycle.
Urea cycle colored.
Related Research Articles
The citric acid cycle —also known as the Krebs cycle, Szent-Györgyi-Krebs cycle or the TCA cycle (tricarboxylic acid cycle)—is a series of chemical reactions to release stored energy through the oxidation of acetyl-CoA derived from carbohydrates, fats, and proteins. The Krebs cycle is used by organisms that respire (as opposed to organisms that ferment) to generate energy, either by anaerobic respiration or aerobic respiration. In addition, the cycle provides precursors of certain amino acids, as well as the reducing agent NADH, that are used in numerous other reactions. Its central importance to many biochemical pathways suggests that it was one of the earliest components of metabolism. Even though it is branded as a 'cycle', it is not necessary for metabolites to follow only one specific route; at least three alternative segments of the citric acid cycle have been recognized.
Ornithine is a non-proteinogenic amino acid that plays a role in the urea cycle. Ornithine is abnormally accumulated in the body in ornithine transcarbamylase deficiency. The radical is ornithyl.
Ornithine transcarbamylase (OTC) is an enzyme that catalyzes the reaction between carbamoyl phosphate (CP) and ornithine (Orn) to form citrulline (Cit) and phosphate (Pi). There are two classes of OTC: anabolic and catabolic. This article focuses on anabolic OTC. Anabolic OTC facilitates the sixth step in the biosynthesis of the amino acid arginine in prokaryotes. In contrast, mammalian OTC plays an essential role in the urea cycle, the purpose of which is to capture toxic ammonia and transform it into urea, a less toxic nitrogen source, for excretion.
Carbamoyl phosphate is an anion of biochemical significance. In land-dwelling animals, it is an intermediary metabolite in nitrogen disposal through the urea cycle and the synthesis of pyrimidines. Its enzymatic counterpart, carbamoyl phosphate synthetase I, interacts with a class of molecules called sirtuins, NAD dependent protein deacetylases, and ATP to form carbamoyl phosphate. CP then enters the urea cycle in which it reacts with ornithine to form citrulline.
Hyperammonemia is a metabolic disturbance characterised by an excess of ammonia in the blood. It is a dangerous condition that may lead to brain injury and death. It may be primary or secondary.
Ornithine transcarbamylase deficiency also known as OTC deficiency is the most common urea cycle disorder in humans. Ornithine transcarbamylase, the defective enzyme in this disorder, is the final enzyme in the proximal portion of the urea cycle, responsible for converting carbamoyl phosphate and ornithine into citrulline. OTC deficiency is inherited in an X-linked recessive manner, meaning males are more commonly affected than females.
Oxaloacetic acid (also known as oxalacetic acid or OAA) is a crystalline organic compound with the chemical formula HO2CC(O)CH2CO2H. Oxaloacetic acid, in the form of its conjugate base oxaloacetate, is a metabolic intermediate in many processes that occur in animals. It takes part in gluconeogenesis, the urea cycle, the glyoxylate cycle, amino acid synthesis, fatty acid synthesis and the citric acid cycle.
In the mitochondrion, the matrix is the space within the inner membrane. The word "matrix" stems from the fact that this space is viscous, compared to the relatively aqueous cytoplasm. The mitochondrial matrix contains the mitochondrial DNA, ribosomes, soluble enzymes, small organic molecules, nucleotide cofactors, and inorganic ions.[1] The enzymes in the matrix facilitate reactions responsible for the production of ATP, such as the citric acid cycle, oxidative phosphorylation, oxidation of pyruvate, and the beta oxidation of fatty acids.
Citrullinemia is an autosomal recessive urea cycle disorder that causes ammonia and other toxic substances to accumulate in the blood.
Argininosuccinate synthase or synthetase is an enzyme that catalyzes the synthesis of argininosuccinate from citrulline and aspartate. In humans, argininosuccinate synthase is encoded by the ASS gene located on chromosome 9.
The enzyme argininosuccinate lyase (EC 4.3.2.1, ASL, argininosuccinase; systematic name 2-(N ω-L-arginino)succinate arginine-lyase (fumarate-forming)) catalyzes the reversible breakdown of argininosuccinate:
N-Acetylglutamate synthase (NAGS) is an enzyme that catalyses the production of N-acetylglutamate (NAG) from glutamate and acetyl-CoA.
Carbamoyl phosphate synthetase I (CPS I) is a ligase enzyme located in the mitochondria involved in the production of urea. Carbamoyl phosphate synthetase I (CPS1 or CPSI) transfers an ammonia molecule to a molecule of bicarbonate that has been phosphorylated by a molecule of ATP. The resulting carbamate is then phosphorylated with another molecule of ATP. The resulting molecule of carbamoyl phosphate leaves the enzyme.
N-Acetylglutamate synthase deficiency is an autosomal recessive urea cycle disorder.
Carbamoyl phosphate synthetase I deficiency is an autosomal recessive metabolic disorder that causes ammonia to accumulate in the blood due to a lack of the enzyme carbamoyl phosphate synthetase I. Ammonia, which is formed when proteins are broken down in the body, is toxic if the levels become too high. The nervous system is especially sensitive to the effects of excess ammonia.
Carbamoyl phosphate synthetase catalyzes the ATP-dependent synthesis of carbamoyl phosphate from glutamine or ammonia and bicarbonate. This enzyme catalyzes the reaction of ATP and bicarbonate to produce carboxy phosphate and ADP. Carboxy phosphate reacts with ammonia to give carbamic acid. In turn, carbamic acid reacts with a second ATP to give carbamoyl phosphate plus ADP.
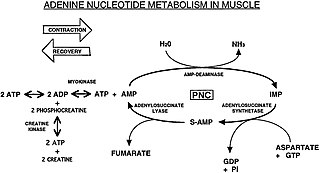
The Purine Nucleotide Cycle is a metabolic pathway in protein metabolism requiring the amino acids aspartate and glutamate. The cycle is used to regulate the levels of adenine nucleotides, in which ammonia and fumarate are generated. AMP coverts into IMP and the byproduct ammonia. IMP converts to S-AMP (adenylosuccinate), which then coverts to AMP and the byproduct fumarate. The fumarate goes on to produce ATP (energy) via oxidative phosphorylation as it enters the Krebs cycle and then the electron transport chain. Lowenstein first described this pathway and outlined its importance in processes including amino acid catabolism and regulation of flux through glycolysis and the Krebs cycle.
Arginine and proline metabolism is one of the central pathways for the biosynthesis of the amino acids arginine and proline from glutamate. The pathways linking arginine, glutamate, and proline are bidirectional. Thus, the net utilization or production of these amino acids is highly dependent on cell type and developmental stage. Altered proline metabolism has been linked to metastasis formation in breast cancer.
Citrullinemia type I (CTLN1), also known as arginosuccinate synthetase deficiency, is a rare disease caused by a deficiency in argininosuccinate synthetase, an enzyme involved in excreting excess nitrogen from the body. There are mild and severe forms of the disease, which is one of the urea cycle disorders.
Carbamoyl phosphate synthetase III is one of the three isoforms of the carbamoyl phosphate synthetase, an enzyme that catalyzes the active production of carbamoyl phosphate in many organisms.
1 2 3 4 Tymoczko, John L.; Berg, Jeremy M.; Stryer, Lubert (2013). BIOCHEMISTRY A Short Course. W.H. Freeman and Company, New York. p.529. ISBN978-1-4292-8360-1.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.