Armadillos are New World placental mammals in the order Cingulata. They form part of the superorder Xenarthra, along with the anteaters and sloths. 21 extant species of armadillo have been described, some of which are distinguished by the number of bands on their armor. All species are native to the Americas, where they inhabit a variety of different environments.

Placental mammals are one of the three extant subdivisions of the class Mammalia, the other two being Monotremata and Marsupialia. Placentalia contains the vast majority of extant mammals, which are partly distinguished from monotremes and marsupials in that the fetus is carried in the uterus of its mother to a relatively late stage of development. The name is something of a misnomer considering that marsupials also nourish their fetuses via a placenta, though for a relatively briefer period, giving birth to less developed young which are then nurtured for a period inside the mother's pouch. Placentalia represents the only living group within Eutheria, which contains all mammals more closely related to placentals than to marsupials.

Megatherium is an extinct genus of ground sloths endemic to South America that lived from the Early Pliocene through the end of the Pleistocene. It is best known for the elephant-sized type species M. americanum, sometimes known as the giant ground sloth, or the megathere, native to the Pampas through southern Bolivia during the Pleistocene. Various other smaller species belonging to the subgenus Pseudomegatherium are known from the Andes.

Glyptodonts are an extinct clade of large, heavily armoured armadillos, reaching up to 1.5 metres (4.9 ft) in height, and maximum body masses of around 2 tonnes. They had short, deep skulls, a fused vertebral column, and a large bony carapace made up of hundreds of individual scutes. Some glyptodonts had clubbed tails, similar to ankylosaurid dinosaurs.
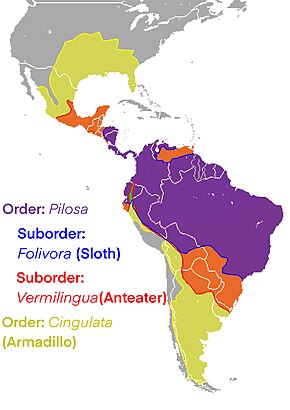
Cingulata, part of the superorder Xenarthra, is an order of armored New World placental mammals. Dasypodids and chlamyphorids, the armadillos, are the only surviving families in the order. Two groups of cingulates much larger than extant armadillos existed until recently: pampatheriids, which reached weights of up to 200 kg (440 lb) and chlamyphorid glyptodonts, which attained masses of 2,000 kg (4,400 lb) or more.

The order Pilosa is a clade of xenarthran placental mammals, native to the Americas. It includes anteaters and sloths. The name comes from the Latin word for "hairy".

Doedicurus is an extinct genus of glyptodont from North and South America containing one species, D. clavicaudatus. Glyptodonts are a member of the family Chlamyphoridae, which also includes some modern armadillo species, and they are classified in the superorder Xenarthra alongside sloths and anteaters. Being a glyptodont, it was a rotund animal with heavy armor and a carapace. Averaging at an approximate 1,400 kg (3,100 lb), it was one of the largest glyptodonts to have ever lived. Though glyptodonts were quadrupeds, large ones like Doedicurus may have been able to stand on two legs like other xenarthrans. It notably sported a spiked tail club, which may have weighed 40 or 65 kg in life, and it may have swung this in defense against predators or in fights with other Doedicurus at speeds of perhaps 11 m/s.

Sloths are a Neotropical group of xenarthran mammals constituting the suborder Folivora, including the extant arboreal tree sloths and extinct terrestrial ground sloths. Noted for their slowness of movement, tree sloths spend most of their lives hanging upside down in the trees of the tropical rainforests of South America and Central America. Sloths are considered to be most closely related to anteaters, together making up the xenarthran order Pilosa.

Eurotamandua is an extinct genus of mammal from extinct family Eurotamanduidae that lived during the middle Eocene.

Tolypeutinae is a subfamily of armadillos in the family Chlamyphoridae, consisting of the giant, three-banded and naked-tailed armadillos.
Glyptodon is a genus of glyptodont, an extinct group of large, herbivorous armadillos, that lived from the Pliocene, around 3.2 million years ago, to the early Holocene, around 11,000 years ago, in Brazil, Uruguay, Paraguay, Bolivia, Peru, Argentina, and Colombia. It is one of, if not the, best known genus of glyptodont. Glyptodon has a long and storied past, being the first named extinct cingulate and the type genus of the subfamily Glyptodontinae. Fossils of Glyptodon have been recorded as early as 1814 from Pleistocene aged deposits from Uruguay, though many were incorrectly referred to the ground sloth Megatherium by early paleontologists.

Euphractinae is an armadillo subfamily in the family Chlamyphoridae.

Holmesina is a genus of pampathere, an extinct group of armadillo-like xenarthrans that were distantly related to extant armadillos. Like armadillos, and unlike the other extinct branch of megafaunal cingulates, the glyptodonts, the shell was made up of flexible plates which allowed the animal to move more easily. Holmesina species were herbivores that grazed on coarse vegetation; armadillos are mostly insectivorous or omnivorous.

Paratheria is an obsolete term for a taxonomic group including the xenarthran mammals and various groups thought to be related to them. It was proposed by Oldfield Thomas in 1887 to set apart the sloths, anteaters, armadillos, and pangolins, usually classified as placentals, from both marsupial and placental mammals, an arrangement that received little support from other workers. When teeth of the extinct gondwanathere mammals were first discovered in Argentina in the 1980s, they were thought to be related to xenarthrans, leading to renewed attention for the hypothesis that xenarthrans are not placentals. However, by the early 1990s, gondwanatheres were shown to be unrelated to xenarthrans, and xenarthrans are still considered to be placentals.

Chlamyphoridae is a family of cingulate mammals. While glyptodonts have traditionally been considered stem-group cingulates outside the group that contains modern armadillos, there had been speculation that the extant family Dasypodidae could be paraphyletic based on morphological evidence. In 2016, an analysis of Doedicurus mtDNA found it was, in fact, nested within the modern armadillos as the sister group of a clade consisting of Chlamyphorinae and Tolypeutinae. For this reason, all extant armadillos but Dasypus were relocated to a new family.

Chlamyphorinae is a subfamily of South American armadillos in the family Chlamyphoridae. Members of this subfamily, the fairy armadillos, are largely fossorial and have reduced eyes and robust forearms with large claws for digging.
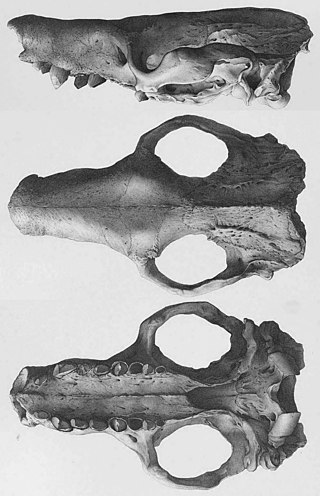
Macroeuphractus is a genus of extinct armadillos from the Late Miocene to Late Pliocene of South America. The genus is noted for its large size, with Macroeuphractus outesi being the largest non-pampathere or glyptodont armadillo discovered, as well as its specializations for carnivory, unique among all xenarthrans.
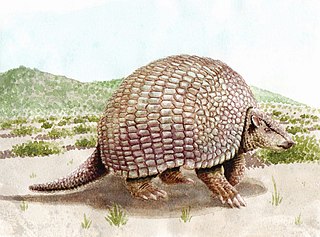
Pampatherium is an extinct genus of xenarthran that lived in the Americas during the Pleistocene. Some species went extinct right at the Pleistocene-Holocene border.