Related Research Articles

The cell nucleus is a membrane-bound organelle found in eukaryotic cells. Eukaryotic cells usually have a single nucleus, but a few cell types, such as mammalian red blood cells, have no nuclei, and a few others including osteoclasts have many. The main structures making up the nucleus are the nuclear envelope, a double membrane that encloses the entire organelle and isolates its contents from the cellular cytoplasm; and the nuclear matrix, a network within the nucleus that adds mechanical support.
In cell biology, an organelle is a specialized subunit, usually within a cell, that has a specific function. The name organelle comes from the idea that these structures are parts of cells, as organs are to the body, hence organelle, the suffix -elle being a diminutive. Organelles are either separately enclosed within their own lipid bilayers or are spatially distinct functional units without a surrounding lipid bilayer. Although most organelles are functional units within cells, some function units that extend outside of cells are often termed organelles, such as cilia, the flagellum and archaellum, and the trichocyst.
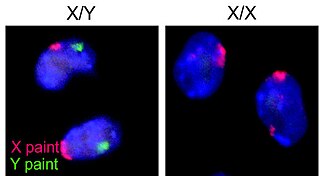
A germ cell is any cell that gives rise to the gametes of an organism that reproduces sexually. In many animals, the germ cells originate in the primitive streak and migrate via the gut of an embryo to the developing gonads. There, they undergo meiosis, followed by cellular differentiation into mature gametes, either eggs or sperm. Unlike animals, plants do not have germ cells designated in early development. Instead, germ cells can arise from somatic cells in the adult, such as the floral meristem of flowering plants.

Spermatogenesis is the process by which haploid spermatozoa develop from germ cells in the seminiferous tubules of the testicle. This process starts with the mitotic division of the stem cells located close to the basement membrane of the tubules. These cells are called spermatogonial stem cells. The mitotic division of these produces two types of cells. Type A cells replenish the stem cells, and type B cells differentiate into primary spermatocytes. The primary spermatocyte divides meiotically into two secondary spermatocytes; each secondary spermatocyte divides into two equal haploid spermatids by Meiosis II. The spermatids are transformed into spermatozoa (sperm) by the process of spermiogenesis. These develop into mature spermatozoa, also known as sperm cells. Thus, the primary spermatocyte gives rise to two cells, the secondary spermatocytes, and the two secondary spermatocytes by their subdivision produce four spermatozoa and four haploid cells.
An oocyte, oöcyte, or ovocyte is a female gametocyte or germ cell involved in reproduction. In other words, it is an immature ovum, or egg cell. An oocyte is produced in a female fetus in the ovary during female gametogenesis. The female germ cells produce a primordial germ cell (PGC), which then undergoes mitosis, forming oogonia. During oogenesis, the oogonia become primary oocytes. An oocyte is a form of genetic material that can be collected for cryoconservation.
In cell biology, a granule is a small particle barely visible by light microscopy. The term is most often used to describe a secretory vesicle containing important components of cell phyisology. Examples of granules include granulocytes, platelet granules, insulin granules, germane granules, starch granules, and stress granules.

The spermatid is the haploid male gametid that results from division of secondary spermatocytes. As a result of meiosis, each spermatid contains only half of the genetic material present in the original primary spermatocyte.

Spermatocytes are a type of male gametocyte in animals. They derive from immature germ cells called spermatogonia. They are found in the testis, in a structure known as the seminiferous tubules. There are two types of spermatocytes, primary and secondary spermatocytes. Primary and secondary spermatocytes are formed through the process of spermatocytogenesis.
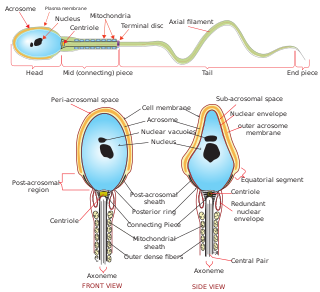
Sperm is the male reproductive cell, or gamete, in anisogamous forms of sexual reproduction. Animals produce motile sperm with a tail known as a flagellum, which are known as spermatozoa, while some red algae and fungi produce non-motile sperm cells, known as spermatia. Flowering plants contain non-motile sperm inside pollen, while some more basal plants like ferns and some gymnosperms have motile sperm.

Spermiogenesis is the final stage of spermatogenesis, during which the spermatids develop into mature spermatozoa. At the beginning of the stage, the spermatid is a more or less circular cell containing a nucleus, Golgi apparatus, centriole and mitochondria; by the end of the process, it has radically transformed into an elongated spermatozoon, complete with a head, midpiece, and tail.
In cellular biology, P-bodies, or processing bodies, are distinct foci formed by phase separation within the cytoplasm of a eukaryotic cell consisting of many enzymes involved in mRNA turnover. P-bodies are highly conserved structures and have been observed in somatic cells originating from vertebrates and invertebrates, plants and yeast. To date, P-bodies have been demonstrated to play fundamental roles in general mRNA decay, nonsense-mediated mRNA decay, adenylate-uridylate-rich element mediated mRNA decay, and microRNA (miRNA) induced mRNA silencing. Not all mRNAs which enter P-bodies are degraded, as it has been demonstrated that some mRNAs can exit P-bodies and re-initiate translation. Purification and sequencing of the mRNA from purified processing bodies showed that these mRNAs are largely translationally repressed upstream of translation initiation and are protected from 5' mRNA decay.
Piwi-interacting RNA (piRNA) is the largest class of small non-coding RNA molecules expressed in animal cells. piRNAs form RNA-protein complexes through interactions with piwi-subfamily Argonaute proteins. These piRNA complexes are mostly involved in the epigenetic and post-transcriptional silencing of transposable elements and other spurious or repeat-derived transcripts, but can also be involved in the regulation of other genetic elements in germ line cells.
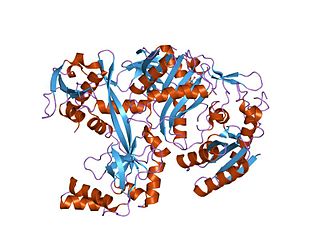
Piwi genes were identified as regulatory proteins responsible for stem cell and germ cell differentiation. Piwi is an abbreviation of P-elementInduced WImpy testis in Drosophila. Piwi proteins are highly conserved RNA-binding proteins and are present in both plants and animals. Piwi proteins belong to the Argonaute/Piwi family and have been classified as nuclear proteins. Studies on Drosophila have also indicated that Piwi proteins have no slicer activity conferred by the presence of the Piwi domain. In addition, Piwi associates with heterochromatin protein 1, an epigenetic modifier, and piRNA-complementary sequences. These are indications of the role Piwi plays in epigenetic regulation. Piwi proteins are also thought to control the biogenesis of piRNA as many Piwi-like proteins contain slicer activity which would allow Piwi proteins to process precursor piRNA into mature piRNA.
Nuage are Drosophila melanogaster germline granules. Nuage are the hallmark of Drosophila melanogaster germline cells, which have an electron-dense perinuclear structure and can silence the selfish genetic elements in Drosophila melanogaster. The term 'Nuage' comes from the French word for 'cloud', as they appear as nebulous electron-dense bodies by electron microscopy. They are found in nurse cells of the developing Drosophila melanogaster egg chamber and are composed of various types of proteins, including RNA-helicases, Tudor domain proteins, Piwi-clade Argonaute proteins, in addition to a PRMT5 methylosome composed of Capsuléen and its co-factor, Valois (MEP50).
Vasa is an RNA binding protein with an ATP-dependent RNA helicase that is a member of the DEAD box family of proteins. The vasa gene is essential for germ cell development and was first identified in Drosophila melanogaster, but has since been found to be conserved in a variety of vertebrates and invertebrates including humans. The Vasa protein is found primarily in germ cells in embryos and adults, where it is involved in germ cell determination and function, as well as in multipotent stem cells, where its exact function is unknown.
The gene Maelstrom, Mael, creates a protein, which was first located in Drosophila melanogaster in the nuage perinuclear structure and has functionality analogous to the spindle, spn, gene class. Its mammalian homolog is MAEL.
Messenger RNP is mRNA with bound proteins. mRNA does not exist "naked" in vivo but is always bound by various proteins while being synthesized, spliced, exported, and translated in the cytoplasm.

The fusome is a membranous structure found in the developing germ cell cysts of many insect orders. Initial description of the fusome occurred in the 19th century and since then the fusome has been extensively studied in Drosophila melanogaster male and female germline development. This structure has roles in maintaining germline cysts, coordinating the number of mitotic divisions prior to meiosis, and oocyte determination by serving as a structure for intercellular communication.
The DAZprotein family is a group of three highly conserved RNA-binding proteins that are important in gametogenesis and meiosis. Therefore, mutations in the genes that encode for the DAZ proteins can have detrimental consequences for fertility.
The germ cell nest forms in the ovaries during their development. The nest consists of multiple interconnected oogonia formed by incomplete cell division. The interconnected oogonia are surrounded by somatic cells called granulosa cells. Later on in development, the germ cell nests break down through invasion of granulosa cells. The result is individual oogonia surrounded by a single layer of granulosa cells. There is also a comparative germ cell nest structure in the developing spermatogonia, with interconnected intracellular cytoplasmic bridges.
References
- ↑ Fawcett, D.; E. Eddy; M. Phillips (1970). "Observations on the fine structure and relationships of the chromatoid body in mammalian spermatogenesis". Biology of Reproduction. 2 (1): 129–153. doi: 10.1095/biolreprod2.1.129 . PMID 4106274.
- 1 2 3 4 5 6 7 Meikar, Oliver; Da Ros, Matteo; Korhonen, Hanna; Kotaja, Noora (August 2011). "Chromatoid body and small RNAs in male germ cells". Reproduction. 142 (2): 195–209. doi: 10.1530/REP-11-0057 . ISSN 1470-1626. PMID 21652638.
- ↑ Anderson, P.; N. Kedersha (2009). "RNA granules: post-transcriptional and epigenetic modulators of gene expression". Nature Reviews Molecular Cell Biology. 10 (6): 430–436. doi:10.1038/nrm2694. PMID 19461665. S2CID 26578027.