Meiosis (; from Ancient Greek μείωσις 'lessening', is a special type of cell division of germ cells in sexually-reproducing organisms that produces the gametes, the sperm or egg cells. It involves two rounds of division that ultimately result in four cells, each with only one copy of each chromosome. Additionally, prior to the division, genetic material from the paternal and maternal copies of each chromosome is crossed over, creating new combinations of code on each chromosome. Later on, during fertilisation, the haploid cells produced by meiosis from a male and a female will fuse to create a zygote, a cell with two copies of each chromosome again.

A spermatozoon is a motile sperm cell produced by male animals relying on internal fertilization. A spermatozoon is a moving form of the haploid cell that is the male gamete that joins with an ovum to form a zygote.
Interphase is the active portion of the cell cycle that includes the G1, S, and G2 phases, where the cell grows, replicates its DNA, and prepares for mitosis, respectively. Interphase was formerly called the "resting phase," but the cell in interphase is not simply dormant. Calling it so would be misleading since a cell in interphase is very busy synthesizing proteins, transcribing DNA into RNA, engulfing extracellular material, and processing signals, to name just a few activities. The cell is quiescent only in G0. Interphase is the phase of the cell cycle in which a typical cell spends most of its life. Interphase is the "daily living" or metabolic phase of the cell, in which the cell obtains nutrients and metabolizes them, grows, replicates its DNA in preparation for mitosis, and conducts other "normal" cell functions.
Gametogenesis is a biological process by which diploid or haploid precursor cells undergo cell division and differentiation to form mature haploid gametes. Depending on the biological life cycle of the organism, gametogenesis occurs by meiotic division of diploid gametocytes into various gametes, or by mitosis. For example, plants produce gametes through mitosis in gametophytes. The gametophytes grow from haploid spores after sporic meiosis. The existence of a multicellular, haploid phase in the life cycle between meiosis and gametogenesis is also referred to as alternation of generations.
A germ cell is any cell that gives rise to the gametes of an organism that reproduces sexually. In many animals, the germ cells originate in the primitive streak and migrate via the gut of an embryo to the developing gonads. There, they undergo meiosis, followed by cellular differentiation into mature gametes, either eggs or sperm. Unlike animals, plants do not have germ cells designated in early development. Instead, germ cells can arise from somatic cells in the adult, such as the floral meristem of flowering plants.
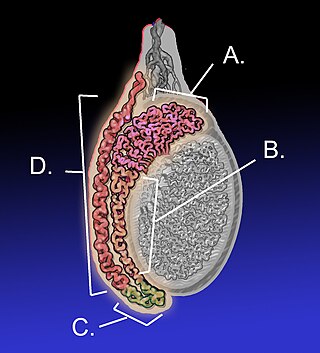
The epididymis is an elongated tubular genital organ attached to the posterior side of each one of the two male reproductive glands, the testicles. It is a single, narrow, tightly coiled tube in adult humans, 6 to 7 centimetres in length; uncoiled the tube would be approximately 6 m long. It connects the testicle to the vas deferens in the male reproductive system. The epididymis serves as an interconnection between the multiple efferent ducts at the rear of a testicle (proximally), and the vas deferens (distally). Its primary function is the storage, maturation and transport of sperm cells.

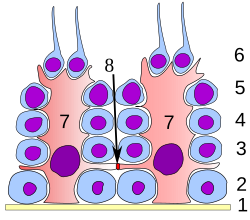
Spermatogenesis is the process by which haploid spermatozoa develop from germ cells in the seminiferous tubules of the testicle. This process starts with the mitotic division of the stem cells located close to the basement membrane of the tubules. These cells are called spermatogonial stem cells. The mitotic division of these produces two types of cells. Type A cells replenish the stem cells, and type B cells differentiate into primary spermatocytes. The primary spermatocyte divides meiotically into two secondary spermatocytes; each secondary spermatocyte divides into two equal haploid spermatids by Meiosis II. The spermatids are transformed into spermatozoa (sperm) by the process of spermiogenesis. These develop into mature spermatozoa, also known as sperm cells. Thus, the primary spermatocyte gives rise to two cells, the secondary spermatocytes, and the two secondary spermatocytes by their subdivision produce four spermatozoa and four haploid cells.
An oocyte, oöcyte, or ovocyte is a female gametocyte or germ cell involved in reproduction. In other words, it is an immature ovum, or egg cell. An oocyte is produced in a female fetus in the ovary during female gametogenesis. The female germ cells produce a primordial germ cell (PGC), which then undergoes mitosis, forming oogonia. During oogenesis, the oogonia become primary oocytes. An oocyte is a form of genetic material that can be collected for cryoconservation.

Non-homologous end joining (NHEJ) is a pathway that repairs double-strand breaks in DNA. It is called "non-homologous" because the break ends are directly ligated without the need for a homologous template, in contrast to homology directed repair (HDR), which requires a homologous sequence to guide repair. NHEJ is active in both non-dividing and proliferating cells, while HDR is not readily accessible in non-dividing cells. The term "non-homologous end joining" was coined in 1996 by Moore and Haber.

Spermatocytes are a type of male gametocyte in animals. They derive from immature germ cells called spermatogonia. They are found in the testis, in a structure known as the seminiferous tubules. There are two types of spermatocytes, primary and secondary spermatocytes. Primary and secondary spermatocytes are formed through the process of spermatocytogenesis.
Reproductive biology includes both sexual and asexual reproduction.
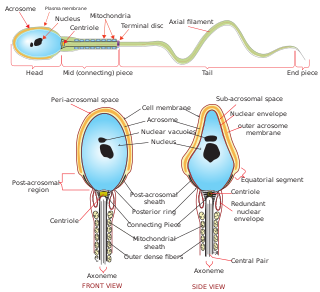
Sperm is the male reproductive cell, or gamete, in anisogamous forms of sexual reproduction. Animals produce motile sperm with a tail known as a flagellum, which are known as spermatozoa, while some red algae and fungi produce non-motile sperm cells, known as spermatia. Flowering plants contain non-motile sperm inside pollen, while some more basal plants like ferns and some gymnosperms have motile sperm.

Spermiogenesis is the final stage of spermatogenesis, during which the spermatids develop into mature spermatozoa. At the beginning of the stage, the spermatid is a more or less circular cell containing a nucleus, Golgi apparatus, centriole and mitochondria; by the end of the process, it has radically transformed into an elongated spermatozoon, complete with a head, midpiece, and tail.

Spermatidogenesis is the creation of spermatids from secondary spermatocytes during spermatogenesis.

Spermatid nuclear transition protein 1 is a protein that in humans is encoded by the TNP1 gene.

Nuclear transition protein 2 is a protein that in humans is encoded by the TNP2 gene.
Spermatozoa develop in the seminiferous tubules of the testes. During their development, the spermatogonia proceed through meiosis to become spermatozoa. Many changes occur during this process: the DNA in nuclei becomes condensed; the acrosome develops as a structure close to the nucleus. The acrosome is derived from the Golgi apparatus and contains hydrolytic enzymes important for fusion of the spermatozoon with an egg cell. During spermiogenesis, the nucleus condenses and changes shape. Abnormal shape change is a feature of sperm in male infertility. The acroplaxome is a structure found between the acrosomal membrane and the nuclear membrane. The acroplaxome contains structural proteins including keratin 5, F-actin and profilin IV.
The leptotene stage, also known as leptonema, is the first of five substages of prophase I during meiosis, the specialized cell division that reduces the chromosome number by half to produce haploid gametes in sexually reproducing organisms.

A spermatogonial stem cell (SSC), also known as a type A spermatogonium, is a spermatogonium that does not differentiate into a spermatocyte, a precursor of sperm cells. Instead, they continue dividing into other spermatogonia or remain dormant to maintain a reserve of spermatogonia. Type B spermatogonia, on the other hand, differentiate into spermatocytes, which in turn undergo meiosis to eventually form mature sperm cells.

Sperm Chromatin Structure Assay (SCSA) is a diagnostic approach that detects sperm abnormality with a large extent of DNA fragmentation. First described by Evenson in 1980, the assay is a flow cytometric test that detects the vulnerability of sperm DNA to acid-induced denaturation DNA in situ. SCSA measures sperm DNA fragmentation attributed to intrinsic and extrinsic factors and reports the degree of fragmentation in terms of DNA Fragmentation Index (DFI). The use of SCSA expands from evaluation of male infertility and subfertility, toxicology studies and evaluation of quality of laboratory semen samples. Notably, SCSA outcompetes other convention sperm DNA fragmentation (sDF) assays such as TUNEL and COMET in terms of efficiency, objectivity, and repeatability.