
Self-pollination is a form of pollination in which pollen from the same plant arrives at the stigma of a flower or at the ovule. There are two types of self-pollination: in autogamy, pollen is transferred to the stigma of the same flower; in geitonogamy, pollen is transferred from the anther of one flower to the stigma of another flower on the same flowering plant, or from microsporangium to ovule within a single (monoecious) gymnosperm. Some plants have mechanisms that ensure autogamy, such as flowers that do not open (cleistogamy), or stamens that move to come into contact with the stigma. The term selfing that is often used as a synonym, is not limited to self-pollination, but also applies to other types of self-fertilization.

Plant reproductive morphology is the study of the physical form and structure of those parts of plants directly or indirectly concerned with sexual reproduction.
Self-incompatibility (SI) is a general name for several genetic mechanisms that prevent self-fertilization in sexually reproducing organisms, and thus encourage outcrossing and allogamy. It is contrasted with separation of sexes among individuals (dioecy), and their various modes of spatial (herkogamy) and temporally (dichogamy) separation.

Heterostyly is a unique form of polymorphism and herkogamy in flowers. In a heterostylous species, two or three morphological types of flowers, termed "morphs", exist in the population. On each individual plant, all flowers share the same morph. The flower morphs differ in the lengths of the pistil and stamens, and these traits are not continuous. The morph phenotype is genetically linked to genes responsible for a unique system of self-incompatibility, termed heteromorphic self-incompatibility, that is, the pollen from a flower on one morph cannot fertilize another flower of the same morph.
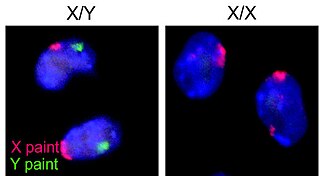
Sequential hermaphroditism is a type of hermaphroditism that occurs in many fish, gastropods, and plants. Sequential hermaphroditism occurs when the individual changes its sex at some point in its life. In particular, a sequential hermaphrodite produces eggs and sperm at different stages in life. Species that can undergo these changes from one sex to another do so as a normal event within their reproductive cycle that is usually cued by either social structure or the achievement of a certain age or size.
Geitonogamy is a type of self-pollination. Geitonogamous pollination is sometimes distinguished from the fertilizations that can result from it, geitonogamy. If a plant is self-incompatible, geitonogamy can reduce seed production.
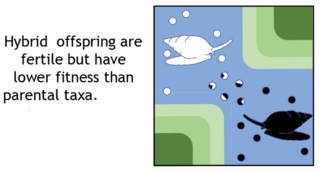
In biology, outbreeding depression happens when crosses between two genetically distant groups or populations result in a reduction of fitness. This is particularly likely if the subspecies have different habitats or if no genetic exchange has occurred, except in the distant past. The concept is in contrast to inbreeding depression, although the two effects can occur simultaneously. The risks of outbreeding are on par with the risks of inbreeding, and these risks sometimes limits the potential for genetic rescue or augmentations. Indeed, studies that report hybridization in mammals find resulting negative consequences about 4 times more likely than positive consequences. Outbreeding depression can occur between an invasive population and a native populations; hybridization can result in extinction of the native species or the loss of native adaptations. Outbreeding depression considered post-zygotic response because outbreeding depression is noted usually in the performance of the progeny.
Allogamy or cross-fertilization is the fertilization of an ovum from one individual with the spermatozoa of another. By contrast, autogamy is the term used for self-fertilization. In humans, the fertilization event is an instance of allogamy. Self-fertilization occurs in hermaphroditic organisms where the two gametes fused in fertilization come from the same individual. This is common in plants and certain protozoans.
The mechanisms of reproductive isolation are a collection of evolutionary mechanisms, behaviors and physiological processes critical for speciation. They prevent members of different species from producing offspring, or ensure that any offspring are sterile. These barriers maintain the integrity of a species by reducing gene flow between related species.
Tristyly is a rare floral polymorphism that consists of three floral morphs that differ in regard to the length of the stamens and style within the flower. This type of floral mechanism is thought to encourage outcross pollen transfer and is usually associated with heteromorphic self-incompatibility to reduce inbreeding. It is an example of heterostyly and reciprocal herkogamy, like distyly, which is the more common form of heterostyly. Darwin first described tristylous species in 1877 in terms of the incompatibility of these three morphs.
Flowering synchrony is the amount of overlap between flowering periods of plants in their mating season compared to what would be expected to occur randomly under given environmental conditions. A population which is flowering synchronously has more plants flowering at the same time than would be expected to occur randomly. A population which is flowering asynchronously has fewer plants flowering at the same time than would be expected randomly. Flowering synchrony can describe synchrony of flowering periods within a year, across years, and across species in a community. There are fitness benefits and disadvantages to synchronized flowering, and it is a widespread phenomenon across pollination syndromes.
Autogamy, or self-fertilization, refers to the fusion of two gametes that come from one individual. Autogamy is predominantly observed in the form of self-pollination, a reproductive mechanism employed by many flowering plants. However, species of protists have also been observed using autogamy as a means of reproduction. Flowering plants engage in autogamy regularly, while the protists that engage in autogamy only do so in stressful environments.

The monocots are one of the two major groups of flowering plants, the other being the dicots. In order to reproduce they utilize various strategies such as employing forms of asexual reproduction, restricting which individuals they are sexually compatible with, or influencing how they are pollinated. Nearly all reproductive strategies that evolved in the dicots have independently evolved in monocots as well. Despite these similarities and their close relatedness, monocots and dicots have distinct traits in their reproductive biologies.

A mixed mating system, also known as “variable inbreeding” a characteristic of many hermaphroditic seed plants, where more than one means of mating is used. Mixed mating usually refers to the production of a mixture of self-fertilized (selfed) and outbred (outcrossed) seeds. Plant mating systems influence the distribution of genetic variation within and among populations, by affecting the propensity of individuals to self-fertilize or cross-fertilize . Mixed mating systems are generally characterized by the frequency of selfing vs. outcrossing, but may include the production of asexual seeds through agamospermy. The trade offs for each strategy depend on ecological conditions, pollinator abundance and herbivory and parasite load. Mating systems are not permanent within species; they can vary with environmental factors, and through domestication when plants are bred for commercial agriculture.
Reproductive assurance occurs as plants have mechanisms to assure full seed set through selfing when outcross pollen is limiting. It is assumed that self-pollination is beneficial, in spite of potential fitness costs, when there is insufficient pollinator services or outcross pollen from other individuals to accomplish full seed set.. This phenomenon has been observed since the 19th century, when Darwin observed that self-pollination was common in some plants. Constant pollen limitation may cause the evolution of automatic selfing, also known as autogamy. This occurs in plants such as weeds, and is a form of reproductive assurance. As plants pursue reproductive assurance through self-fertilization, there is an increase in homozygosity, and inbreeding depression, due to genetic load, which results in reduced fitness of selfed offspring. Solely outcrossing plants may not be successful colonizers of new regions due to lack of other plants to outcross with, so colonizing species are expected to have mechanisms of reproductive assurance - an idea first proposed by Herbert Baker and referred to as Baker's Law. Baker’s Law predicts that reproductive assurance should be common in weedy plants that persist by colonizing new sites. As plants evolve towards increase self-fertilization, energy is redirected to seed production rather than characteristics that increased outcrossing, such as floral attractants, which is a condition known as the selfing syndrome.

Floral color change occurs in flowers in a wide range of angiosperm taxa that undergo a color change associated with their age, or after successful pollination.

Distyly is a type of heterostyly in which a plant demonstrates reciprocal herkogamy. This breeding system is characterized by two separate flower morphs, where individual plants produce flowers that either have long styles and short stamens, or that have short styles and long stamens. However, distyly can refer to any plant that shows some degree of self-incompatibility and has two morphs if at least one of the following characteristics is true; there is a difference in style length, filament length, pollen size or shape, or the surface of the stigma. Specifically these plants exhibit intra-morph self-incompatibility, flowers of the same style morph are incompatible. Distylous species that do not exhibit true self-incompatibility generally show a bias towards inter-morph crosses - meaning they exhibit higher success rates when reproducing with an individual of the opposite morph.
Late-acting self-incompatibility (LSI) is the occurrence of self-incompatibility (SI) in flowering plants where pollen tubes from self-pollen successfully reach the ovary, but ovules fail to develop. Mechanisms that might cause late-acting self-incompatibility have yet to be elucidated. One hypothesis is that the occurrence of LSI is caused by early-acting inbreeding depression where the expression of genetic load causes self-fertilized embryos to abort.
Gametophytic selection is the selection of one haploid pollen grain over another through the means of pollen competition, and that resulting sporophytic generations are positively affected by this competition. Evidence for the positive effects of gametophytic selection on the sporophyte generation has been observed in several flowering plant species, but there are is still some debate as to the biological significance of gametophytic selection.
June Nasrallah is Barbara McClintock Professor in the Plant Biology Section of the School of Integrative Plant Science at Cornell University. Her research focuses on plant reproductive biology and the cell-cell interactions that underlie self-incompatibility in plants belonging to the mustard (Brassicaceae) family. She was elected to the US National Academy of Sciences in 2003 for this work and her contributions generally to our understanding of receptor-based signalling in plants.