| Cystine-knot domain | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
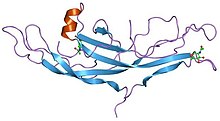 Structure of human chorionic gonadotropin. [1] | |||||||||||
| Identifiers | |||||||||||
| Symbol | Cys_knot | ||||||||||
| Pfam | PF00007 | ||||||||||
| Pfam clan | CL0079 | ||||||||||
| InterPro | IPR006208 | ||||||||||
| SCOP2 | 1hcn / SCOPe / SUPFAM | ||||||||||
| |||||||||||
A cystine knot is a protein structural motif containing three disulfide bridges (formed from pairs of cysteine residues). The sections of polypeptide that occur between two of them form a loop through which a third disulfide bond passes, forming a rotaxane-like substructure. The cystine knot motif stabilizes protein structure and is conserved in proteins across various species. [2] [3] [4] There are three types of cystine knot, which differ in the topology of the disulfide bonds: [5]
- Growth factor cystine knot (GFCK)
- Inhibitor cystine knot (ICK), common in spider and snail toxins
- Cyclic cystine knot, or cyclotide
The growth factor cystine knot was first observed in the structure of nerve growth factor (NGF), solved by X-ray crystallography and published in 1991. [6] The GFCK is present in four superfamilies. These include nerve growth factor, transforming growth factor beta (TGF-β), platelet-derived growth factor, and glycoprotein hormones including human chorionic gonadotropin. These are structurally related due to the presence of the cystine knot motif but differ in sequence. [7] All GFCK structures that have been determined are dimeric, but their dimerization modes in different classes are different. [8] The vascular endothelial growth factor subfamily, categorized as part of the platelet-derived growth factor superfamily, includes proteins that are angiogenic factors. [9]
The presence of the cyclic cystine knot (CCK) motif was discovered when cyclotides were isolated from various plant families. The CCK motif has a cyclic backbone, triple stranded beta sheet, and cystine knot conformation. [10]
Inhibitor cystine knot (ICK) is a structural motif with a triple stranded antiparallel beta sheet linked by three disulfide bonds, forming a knotted core. It is often found in many venom peptides such as those of snails, spiders, and scorpions. Peptide K-PVIIA, which contains an ICK, can undergo a successful enzymatic backbone cyclization. The disulfide connectivity and the common sequence pattern of the ICK motif provides the stability of the peptides that support cyclization. [11]