
Disc shedding is the process by which photoreceptor cells in the retina are renewed. The disc formations in the outer segment of photoreceptors, which contain the photosensitive opsins, are completely renewed every ten days.
Disc shedding is the process by which photoreceptor cells in the retina are renewed. The disc formations in the outer segment of photoreceptors, which contain the photosensitive opsins, are completely renewed every ten days.
The retina contains two types of photoreceptor – rod cells and cone cells. There are about 6-7 million cones that mediate photopic vision, and they are concentrated in the macula at the center of the retina. There are about 120 million rods that are more sensitive than the cones and therefore mediate scotopic vision.
A vertebrate's photoreceptors are divided into three parts:
The photosensitive outer segment consists of a series of discrete membranous discs . [1]
While in the rod, these discs lack any direct connection to the surface membrane (with the exception of a few recently formed basal discs that remain in continuity with the surface), the cone's photosensitive membrane is continuous with the surface membrane. The outer segment (OS) discs are densely packed with rhodopsin for high-sensitivity light detection. [2] These discs are completely replaced once every ten days and this continuous renewal continues throughout the lifetime of the sighted animal.
After the opsins are synthesized, they fuse to the plasma membrane, which then invaginates with discs budding off internally, forming the tightly packed stacks of outer segment discs. From translation of opsin to formation of the discs takes just a couple of hours.
Disc shedding was first described by RW Young in 1967. [3] Discs mature along with their distal migration; aged discs shed at the distal tip and are engulfed by the neighboring retinal pigment epithelial (RPE) cells for degradation. [2]
One early study showed that cones may not experience the cone shedding as rods do and may renew by replacing molecular constituents individually. [3] However, other studies do show that at least some mammalian cones do shed their discs as a normal ongoing process. [4]
Each day about one tenth of the length of the outer segment is lost, so that after ten days the entire outer segment has been replaced. Regulating factors are involved at each step. While disc assembly is mostly genetically controlled, disc shedding and the subsequent RPE phagocytosis appear to be regulated by environmental factors like light and temperature. [5]
The timing of shedding follows a circadian rhythm according to neuromodulators, namely dopamine and melatonin. Melatonin is synthesized by the photoreceptors at night, and is inhibited by light and dopamine, so triggers cone disc shedding. Dopamine production is stimulated by light and inhibited by dark and melatonin, so triggers cone disc shedding. Importantly, rod discs are shed during the day and cone discs are shed during the night. [6]
One grey area in the entire mechanism of outer segment disc shedding is in what exactly triggers the detachment of the discs and how they are transported out of the OS and phagocytosed by the RPE cells.
Some studies suggest that disc detachment precedes engulfment by the RPE cells, and that an active process in the rod outer segment severs the disc. [3] [4] However, other studies observed RPE cell processes intruding into the OS during disc detachment. These processes are structurally similar to processes formed by macrophages during phagocytosis and were accordingly referred to as pseudopodia. The study suggested that these pseudopodia were the organelles of phagocytosis and that they may play a direct role in disc detachment. [7]
A 2007 paper offers a third theory that builds on recent evidence that suggests that rhodopsin-deficient mice fail to develop OSS. [8] [9] Researchers at Cornell hypothesized that rhodopsin itself has a role in OS biogenesis, in addition to its role as a phototransduction receptor. [2] While the molecular basis underlying rhodopsin's participation in OS development is unknown, emerging evidence suggests that rhodopsin's cytoplasmic C-terminal tail bears an “address signal” for its transport from its site of synthesis in the rod cell body to the OS. [10] [11]

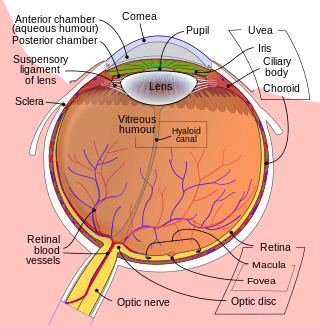
The retina is the innermost, light-sensitive layer of tissue of the eye of most vertebrates and some molluscs. The optics of the eye create a focused two-dimensional image of the visual world on the retina, which then processes that image within the retina and sends nerve impulses along the optic nerve to the visual cortex to create visual perception. The retina serves a function which is in many ways analogous to that of the film or image sensor in a camera.

Retinitis pigmentosa (RP) is a genetic disorder of the eyes that causes loss of vision. Symptoms include trouble seeing at night and decreasing peripheral vision. As peripheral vision worsens, people may experience "tunnel vision". Complete blindness is uncommon. Onset of symptoms is generally gradual and often begins in childhood.

A photoreceptor cell is a specialized type of neuroepithelial cell found in the retina that is capable of visual phototransduction. The great biological importance of photoreceptors is that they convert light into signals that can stimulate biological processes. To be more specific, photoreceptor proteins in the cell absorb photons, triggering a change in the cell's membrane potential.

Rod cells are photoreceptor cells in the retina of the eye that can function in lower light better than the other type of visual photoreceptor, cone cells. Rods are usually found concentrated at the outer edges of the retina and are used in peripheral vision. On average, there are approximately 92 million rod cells in the human retina. Rod cells are more sensitive than cone cells and are almost entirely responsible for night vision. However, rods have little role in color vision, which is the main reason why colors are much less apparent in dim light.
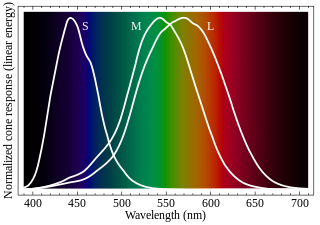
Cone cells, or cones, are photoreceptor cells in the retinas of vertebrates' eyes, including the human eye. They respond differently to light of different wavelengths, and the combination of their responses is responsible for color vision. Cones function best in relatively bright light, called the photopic region, as opposed to rod cells, which work better in dim light, or the scotopic region. Cone cells are densely packed in the fovea centralis, a 0.3 mm diameter rod-free area with very thin, densely packed cones which quickly reduce in number towards the periphery of the retina. Conversely, they are absent from the optic disc, contributing to the blind spot. There are about six to seven million cones in a human eye, with the highest concentration being towards the macula.
In visual physiology, adaptation is the ability of the retina of the eye to adjust to various levels of light. Natural night vision, or scotopic vision, is the ability to see under low-light conditions. In humans, rod cells are exclusively responsible for night vision as cone cells are only able to function at higher illumination levels. Night vision is of lower quality than day vision because it is limited in resolution and colors cannot be discerned; only shades of gray are seen. In order for humans to transition from day to night vision they must undergo a dark adaptation period of up to two hours in which each eye adjusts from a high to a low luminescence "setting", increasing sensitivity hugely, by many orders of magnitude. This adaptation period is different between rod and cone cells and results from the regeneration of photopigments to increase retinal sensitivity. Light adaptation, in contrast, works very quickly, within seconds.

Melanopsin is a type of photopigment belonging to a larger family of light-sensitive retinal proteins called opsins and encoded by the gene Opn4. In the mammalian retina, there are two additional categories of opsins, both involved in the formation of visual images: rhodopsin and photopsin in the rod and cone photoreceptor cells, respectively.
Photopigments are unstable pigments that undergo a chemical change when they absorb light. The term is generally applied to the non-protein chromophore moiety of photosensitive chromoproteins, such as the pigments involved in photosynthesis and photoreception. In medical terminology, "photopigment" commonly refers to the photoreceptor proteins of the retina.
Visual phototransduction is the sensory transduction process of the visual system by which light is detected to yield nerve impulses in the rod cells and cone cells in the retina of the eye in humans and other vertebrates. It relies on the visual cycle, a sequence of biochemical reactions in which a molecule of retinal bound to opsin undergoes photoisomerization, initiates a cascade that signals detection of the photon, and is indirectly restored to its photosensitive isomer for reuse. Phototransduction in some invertebrates such as fruit flies relies on similar processes.
Intrinsically photosensitive retinal ganglion cells (ipRGCs), also called photosensitive retinal ganglion cells (pRGC), or melanopsin-containing retinal ganglion cells (mRGCs), are a type of neuron in the retina of the mammalian eye. The presence of ipRGCs was first suspected in 1927 when rodless, coneless mice still responded to a light stimulus through pupil constriction, This implied that rods and cones are not the only light-sensitive neurons in the retina. Yet research on these cells did not advance until the 1980s. Recent research has shown that these retinal ganglion cells, unlike other retinal ganglion cells, are intrinsically photosensitive due to the presence of melanopsin, a light-sensitive protein. Therefore they constitute a third class of photoreceptors, in addition to rod and cone cells.

The pigmented layer of retina or retinal pigment epithelium (RPE) is the pigmented cell layer just outside the neurosensory retina that nourishes retinal visual cells, and is firmly attached to the underlying choroid and overlying retinal visual cells.
Ignacio Provencio is an American neuroscientist and the discoverer of melanopsin, an opsin found in specialized photosensitive ganglion cells of the mammalian retina. Provencio served as the program committee chair of the Society for Research on Biological Rhythms from 2008 to 2010.
Rhodopsin kinase is a serine/threonine-specific protein kinase involved in phototransduction. This enzyme catalyses the following chemical reaction:
The visual cycle is a process in the retina that replenishes the molecule retinal for its use in vision. Retinal is the chromophore of most visual opsins, meaning it captures the photons to begin the phototransduction cascade. When the photon is absorbed, the 11-cis retinal photoisomerizes into all-trans retinal as it is ejected from the opsin protein. Each molecule of retinal must travel from the photoreceptor cell to the RPE and back in order to be refreshed and combined with another opsin. This closed enzymatic pathway of 11-cis retinal is sometimes called Wald's visual cycle after George Wald (1906–1997), who received the Nobel Prize in 1967 for his work towards its discovery.
Mammals normally have a pair of eyes. Although mammalian vision is not so excellent as bird vision, it is at least dichromatic for most of mammalian species, with certain families possessing a trichromatic color perception.

Retinal degeneration is a retinopathy which consists in the deterioration of the retina caused by the progressive death of its cells. There are several reasons for retinal degeneration, including artery or vein occlusion, diabetic retinopathy, R.L.F./R.O.P., or disease. These may present in many different ways such as impaired vision, night blindness, retinal detachment, light sensitivity, tunnel vision, and loss of peripheral vision to total loss of vision. Of the retinal degenerative diseases retinitis pigmentosa (RP) is a very important example.
Retinal gene therapy holds a promise in treating different forms of non-inherited and inherited blindness.

Emixustat is a small molecule notable for its establishment of a new class of compounds known as visual cycle modulators (VCMs). Formulated as the hydrochloride salt, emixustat hydrochloride, it is the first synthetic medicinal compound shown to affect retinal disease processes when taken by mouth. Emixustat was invented by the British-American chemist, Ian L. Scott, and is currently in Phase 3 trials for dry, age-related macular degeneration (AMD).

William Archer Hagins was an American medical researcher. He was chief of the Section of Membrane Biophysics in National Institute of Diabetes and Digestive and Kidney Diseases's Laboratory of Chemical Physics upon his retirement in 2007. Hagins and colleagues made the seminal discovery of the dark current in photoreceptor cells. This finding became central to understanding how the visual cells worked and led to knowledge of the importance of reattaching a detached retina as soon as possible for continued use. As a fellow of Fulbright Program, he'd also served in the United States Navy as a Research Medical Officer. He joined NIDDK's Laboratory of Physical Biology in 1958, doing independent research in the Section of Photobiology, headed by Frederick Sumner Brackett. Hagins was a mentor to many, particularly through his work with the Brackett Foundation.
Vertebrate visual opsins are a subclass of ciliary opsins and mediate vision in vertebrates. They include the opsins in human rod and cone cells. They are often abbreviated to opsin, as they were the first opsins discovered and are still the most widely studied opsins.