| Palpata | |
|---|---|
 | |
| Illustration by Ernst Haeckel | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Annelida |
| Class: | Polychaeta |
| Subclass: | Palpata |
| Orders | |
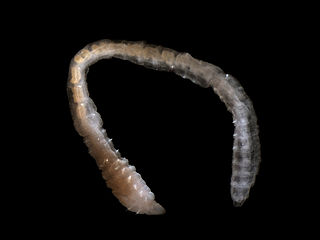
Palpata is a subclass of polychaete worm. Members of this subclass are mostly deposit feeders on marine detritus or filter feeders. Palpata has become superfluous with the elevation of Canalipalpata to subclass. [1]
In biological classification, class is a taxonomic rank, as well as a taxonomic unit, a taxon, in that rank. Other well-known ranks in descending order of size are life, domain, kingdom, phylum, order, family, genus, and species, with class fitting between phylum and order.

{{Paraphyletic group Support and Locomotion Polychaeta. Polychaetes provide a classic example of the employment of coelomic spaces as a hydrostatic skeleton for body support. Coupled with the well developed musculature, the metameric body plan, and the parapodia, this hydrostatic quality provides the basis for understanding locomotion in these worms. We begin a survey of locomotor patterns by examining Nereis, an errant, homonomous polychaete. Keep in mind that in such polychaetes the intersegmental septa are functionally complete, and thus the coelomic spaces in each segment can be effectively isolated hydraulically from each other. Modifications on this fundamental arrangement are discussed later. In addition to burrowing, Nereis can engage in three basic epibenthic locomotor patterns: slow crawling, rapid crawling, and rather inefficient swimming. All of these methods of movement depend primarily on the bands of longitudinal muscles, especially the larger dorsolateral bands, and on the parapodial muscles. The circular muscles are relatively thin and serve primarily to maintain adequate hydrostatic pressure within the coelomic compartments. Each method of locomotion in Nereis involves the antagonistic action of the longitudinal muscles on opposite sides of the body in each segment. During movement, the longitudinal muscles on one side of any given segment alternately contract and relax in opposing synchrony with the action of the muscles on the other side of the segment. Thus, the body is thrown into undulations that move in metachronal waves from posterior to anterior. Variations in the length and amplitude of these waves combine with parapodial movements to produce the different patterns of locomotion. The parapodia and their chaetae are extended maximally in a power stroke as they pass along the crest of each metachronal wave. Conversely, the parapodia and chaetae retract in the wave troughs during their recovery stroke. Thus, the parapodia on opposite sides of any given segment are exactly out of phase with one another. When Nereis is crawling slowly, the body is thrown into a high number of metachronal undulations of short wavelength and low amplitude. The extended parapodial chaetae on the wave crests are pushed against the substratum and serve as pivot points as the parapodium engages in its power stroke. As the parapodium moves past the crest, it is retracted and lifted from the substratum as it is brought forward during its recovery stroke. The main pushing force in this sort of movement is provided by the parapodial muscles. During rapid crawling, much of the driving force is provided by the longitudinal body wall muscles in association with the longer wavelength and greater amplitude of the body undulations, which accentuate the power strokes of the parapodia. Nereis can leave the substratum to engage in a rather inefficient swimming behavior. In swimming, the metachronal wavelength and amplitude are even greater than they are in rapid crawling. When watching a nereid swim, however, one gets the impression that the “harder it tries” the less progress it makes, and there is some truth to this. The problem is that, even though the parapodia act as paddles pushing the animal forward on their power strokes, the large metachronal waves continue to move from posterior to anterior and actually create a water current in that same direction; this current tends to push the animal into reverse. The result is that Nereis is able to lift itself off the substratum, but then largely thrashes about in the water. This behavior is used primarily as a short-term mechanism to escape benthic predators rather than as ameans to get from one place to another. With these basic patterns and mechanisms in mind, we consider a few other methods of locomotion in polychaetes. Nephtys superficially resembles Nereis, but its methods of movement are significantly different. Although Nephtys is less efficient than Nereis at slow walking, it is a much better swimmer; it is also capable of effective burrowing in soft substrata. The large, fleshy parapodia serve as paddles, and, when swimming, Nephtys does not produce long, deep metachronal waves. Rather, the faster it swims, the shorter and shallower the waves become, thus eliminating much of the counterproductive force described for Nereis. When initiating burrowing, Nephtys swims head-first into the substratum, anchors the body by extending the chaetae laterally from the buried segments, and then extends the proboscis deeper into the sand. A swimming motion is then employed to burrow deeper into the substratum. In contrast to the above descriptions, scale worms have capitalized on the use of their muscular parapodia as efficient walking devices. The body undulates little if at all, and there is a corresponding reduction in the size of the longitudinal muscle bands and their importance in locomotion. In fact, these worms depend almost entirely on the action of the parapodia for walking; polynoids cannot swim. Many of the highly efficient burrowers have secondarily lost most of the intersegmental septa, or have septa that are perforated. The loss of complete septa means that segments are not of constant volume; in other words, a loss of coelomic fluid from one body region causes a corresponding gain in another. These polychaetes have reduced parapodia. The chaetae, or simply the surface of the expanded portions of the body, serve as anchor points, while the burrow wall provides an antagonistic force resisting the hydraulic pressure. In Polyphysia, peristaltic waves move constricted body regions forward while the anchored parts provide leverage. The constricted areas are reduced both in diameter and in length by simultaneous contraction of both the circular and the longitudinal muscles. Arenicola burrows first embedding and anchoringlongitudinal Arenicola burrows by first embedding and anchoring the anterior body region in the substratum. The anchoring is accomplished by contracting the circular muscles of the posterior portion of the body, thus forcing coelomic fluid anteriorly and causing the first few segments to swell. Then the posterior longitudinal muscles contract, thereby pulling the back of the worm forward. To continue the burrowing, a second phase of activity is undertaken. As the anterior circular muscles contract and the longitudinal bands relax, the posterior edges of each involved segment are protruded as anchor points to prevent backward movement; the proboscis is thrust forward, deepening the burrow. Then the proboscis is retracted, the front end of the body is engorged with fluid, and the entire process is repeated. Different burrowing mechanisms are known among other polychaetes. For example, Glycera, a long, sleek worm, burrows rapidly using its large, muscular proboscis almost exclusively. The proboscis is thrust into the substratum and swelled; then the body is drawn in by contraction of the proboscis muscles. Most tube-dwelling polychaetes are heteronomous and have rather soft bodies and relatively weak muscles. The parapodia are reduced, so the chaetae are used to position and anchor the animal in its tube. Movement within the tube is usually accomplished by slow peristaltic action of the body or by chaetae movements. When the anterior end is extended for feeding, it may be quickly withdrawn by special retractor muscles while the unexposed portion of the body is anchored in the tube. Polychaete tubes provide protection as well as support for these soft-bodied worms, and also keep the animal oriented properly in relation to the substratum. Some polychaetes build tubes composed entirely of their own secretions. Most notable among these tube builders are the serpulids and spirorbids, which construct their tubes of calcium carbonate secreted by a pair of large glands near a fold of the peristomium called the collar. The crystals of calcium carbonate are added to an organic matrix; the mixture is molded to the top of the tube by the collar fold and held in place until it hardens. Some sabellids produce parchment-like or membranous tubes of organic secretions molded by the collar. Others, such as Sabella, mix mucous secretions with size selected particles extracted from feeding currents, then lay down the tube with this material. Numerous other polychaete groups form similar tubes of sediment particles collected in various ways and cemented together with mucus. A few polychaetes are able to excavate burrows by boring into calcareous substrata, such as rocks, coral skeletons or mollusc shells. In extreme situations, the activity of the polychaetes may have deleterious effects on the “host.” For example, species of Polydora (Spionidae) can cause serious damage to commercially raised oysters. Many sedentary polychaetes use modifications of the basic locomotor actions described above to provide means of moving water through their tubes or burrows. Some of these modifications are discussed in the sectionon feeding.

Detritivores, also known as detrivores, detritophages, detritus feeders, or detritus eaters, are heterotrophs that obtain nutrients by consuming detritus. There are many kinds of invertebrates, vertebrates and plants that carry out coprophagy. By doing so, all these detritivores contribute to decomposition and the nutrient cycles. They should be distinguished from other decomposers, such as many species of bacteria, fungi and protists, which are unable to ingest discrete lumps of matter, but instead live by absorbing and metabolizing on a molecular scale. However, the terms detritivore and decomposer are often used interchangeably.