
Sperm thermotaxis is a form of sperm guidance, in which sperm cells (spermatozoa) actively change their swimming direction according to a temperature gradient, swimming up the gradient. Thus far this process has been discovered in mammals only.
Sperm thermotaxis is a form of sperm guidance, in which sperm cells (spermatozoa) actively change their swimming direction according to a temperature gradient, swimming up the gradient. Thus far this process has been discovered in mammals only.
The discovery of mammalian sperm chemotaxis and the realization that it can guide spermatozoa for short distances only, estimated at the order of millimeters, [1] triggered a search for potential long-range guidance mechanisms. The findings that, at least in rabbits [2] and pigs, [3] a temperature difference exists within the oviduct, and that this temperature difference is established at ovulation in rabbits due to a temperature drop in the oviduct near the junction with the uterus, creating a temperature gradient between the sperm storage site and the fertilization site in the oviduct [4] (Figure 1), led to investigation whether mammalian spermatozoa can respond to a temperature gradient by thermotaxis. [5]
Mammalian sperm thermotaxis was, hitherto, demonstrated in three species: humans, rabbits, and mice. [5] [6] This was done by two methods. One involved a Zigmond chamber, modified to make the temperature in each well separately controllable and measurable. A linear temperature gradient was established between the wells and the swimming of spermatozoa in this gradient was analyzed. A small fraction of the spermatozoa (at the order of ~10%), shown to be the capacitated cells, biased their swimming direction according to the gradient, moving towards the warmer temperature. [5] The other method involved two [7] [8] - or three [6] -compartment separation tube placed within a thermoseparation device that maintains a linear temperature gradient. Sperm accumulation at the warmer end of the separation tube was much higher than the accumulation at the same temperature but in the absence of a temperature gradient. [6] This gradient-dependent sperm accumulation was observed over a wide temperature range (29-41 °C). [8]
Since temperature affects almost every process, much attention has been devoted to the question of whether the measurements, mentioned just above, truly demonstrate thermotaxis or whether they reflect another temperature-dependent process. The most pronounced effect of temperature in liquid is convection, which raised the concern that the apparent thermotactic response could have been a reflection of a passive drift in the liquid current or a rheotactic response [9] to the current (rather than to the temperature gradient per se). Another concern was that the temperature could have changed the local pH of the buffer solution in which the spermatozoa are suspended. This could generate a pH gradient along the temperature gradient, and the spermatozoa might have responded to the formed pH gradient by chemotaxis. However, careful experimental examinations of all these possibilities with proper controls demonstrated that the measured responses to temperature are true thermotactic responses and that they are not a reflection of any other temperature-sensitive process, including rheotaxis and chemotaxis. [1] [8]
The behavioral mechanism of sperm thermotaxis has been so far only investigated in human spermatozoa. [10] Like the behavioral mechanisms of bacterial chemotaxis [11] and human sperm chemotaxis, [12] the behavioral mechanism of human sperm thermotaxis appears to be stochastic rather than deterministic. Capacitated human spermatozoa swim in rather straight lines interrupted by turns and brief episodes of hyperactivation. Each such episode results in swimming in a new direction. When the spermatozoa sense a decrease in temperature, the frequency of turns and hyperactivation events increases due to increased flagellar-wave amplitude that results in enhanced side-to-side head displacement. With time, this response undergoes partial adaptation. The opposite happens in response to an increase in temperature. This suggests that when capacitated spermatozoa swim up a temperature gradient, turns are repressed and the spermatozoa continue swimming in the gradient direction. When they happen to swim down the gradient, they turn again and again until their swimming direction is again up the gradient.
The response of spermatozoa to temporal temperature changes even when the temperature is kept constant spatially [10] suggests that, as in the case of human sperm chemotaxis, [12] [13] sperm thermotaxis involves temporal gradient sensing. In other words, spermatozoa apparently compare the temperature (or a temperature-dependent function) between consecutive time points. This, however, does not exclude the occurrence of spatial temperature sensing in addition to temporal sensing. Human spermatozoa can respond thermotactically within a wide temperature range (at least 29–41 °C). [8] Within this range they preferentially accumulate in warmer temperatures rather than at a single specific, preferred temperature. Amazingly, they can sense and thermotactically respond to temperature gradients as low as <0.014 °C/mm. This means that when human spermatozoa swim a distance that equals their body length (~46 µm) they respond to a temperature difference of <0.0006 °C!
The molecular mechanism underlying thermotaxis, in general, and thermosensing with such extreme sensitivity, in particular, is obscure. It is known that, unlike other recognized thermosensors in mammals, the thermosensors for sperm thermotaxis do not seem to be temperature-sensitive ion channels. They are rather opsins, [6] known to be G-protein-coupled receptors that act as photosensors in vision. The opsins are present in spermatozoa at specific sites, which depend on the species and the opsin type. [6] They are involved in sperm thermotaxis via two signaling pathways—a phospholipase C signaling pathway and a cyclic-nucleotide pathway. The former was shown by pharmacological means in human spermatozoa to involve the enzyme phospholipase C, an inositol trisphosphate receptor calcium channel located on internal calcium stores, the calcium channel TRPC3, and intracellular calcium. [6] [7] The latter was hitherto shown to involve phosphodiesterase. [6] Blocking both pathways fully inhibits sperm thermotaxis. [6]
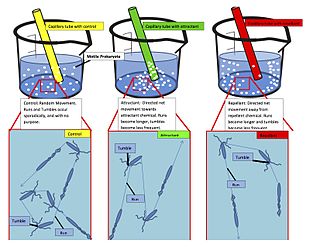
Chemotaxis is the movement of an organism or entity in response to a chemical stimulus. Somatic cells, bacteria, and other single-cell or multicellular organisms direct their movements according to certain chemicals in their environment. This is important for bacteria to find food by swimming toward the highest concentration of food molecules, or to flee from poisons. In multicellular organisms, chemotaxis is critical to early development and development as well as in normal function and health. In addition, it has been recognized that mechanisms that allow chemotaxis in animals can be subverted during cancer metastasis. The aberrant chemotaxis of leukocytes and lymphocytes also contribute to inflammatory diseases such as atherosclerosis, asthma, and arthritis. Sub-cellular components, such as the polarity patch generated by mating yeast, may also display chemotactic behavior.

A spermatozoon is a motile sperm cell, or moving form of the haploid cell that is the male gamete. A spermatozoon joins an ovum to form a zygote.
Fertilisation or fertilization, also known as generative fertilisation, syngamy and impregnation, is the fusion of gametes to give rise to a new individual organism or offspring and initiate its development. While processes such as insemination or pollination which happen before the fusion of gametes are also sometimes informally referred to as fertilisation, these are technically separate processes. The cycle of fertilisation and development of new individuals is called sexual reproduction. During double fertilisation in angiosperms the haploid male gamete combines with two haploid polar nuclei to form a triploid primary endosperm nucleus by the process of vegetative fertilisation.
A taxis is the movement of an organism in response to a stimulus such as light or the presence of food. Taxes are innate behavioural responses. A taxis differs from a tropism in that in the case of taxis, the organism has motility and demonstrates guided movement towards or away from the stimulus source. It is sometimes distinguished from a kinesis, a non-directional change in activity in response to a stimulus.

During fertilization, a sperm must first fuse with the plasma membrane and then penetrate the female egg cell to fertilize it. Fusing to the egg cell usually causes little problem, whereas penetrating through the egg's hard shell or extracellular matrix can be more difficult. Therefore, sperm cells go through a process known as the acrosome reaction, which is the reaction that occurs in the acrosome of the sperm as it approaches the egg.
Thermotaxis is a behavior in which an organism directs its locomotion up or down a gradient of temperature.

Motility is the ability of an organism to move independently, using metabolic energy.
Capacitation is the penultimate step in the maturation of mammalian spermatozoa and is required to render them competent to fertilize an oocyte. This step is a biochemical event; the sperm move normally and look mature prior to capacitation. In vivo, capacitation occurs after ejaculation, when the spermatozoa leave the vagina and enter the upper female reproductive tract. The uterus aids in the steps of capacitation by secreting sterol-binding albumin, lipoproteins, and proteolytic and glycosidasic enzymes such as heparin.
Hyperactivation is a type of sperm motility. Hyperactivated sperm motility is characterised by a high amplitude, asymmetrical beating pattern of the sperm tail (flagellum). This type of motility may aid in sperm penetration of the zona pellucida, which encloses the ovum.
Thermotropism or thermotropic movement is the movement of an organism or a part of an organism in response to heat or changes from the environment's temperature. A common example is the curling of Rhododendron leaves in response to cold temperatures. Mimosa pudica also show thermotropism by the collapsing of leaf petioles leading to the folding of leaflets, when temperature drops.
In biology, polyspermy describes the fertilization of an egg by more than one sperm. Diploid organisms normally contain two copies of each chromosome, one from each parent. The cell resulting from polyspermy, on the other hand, contains three or more copies of each chromosome—one from the egg and one each from multiple sperm. Usually, the result is an unviable zygote. This may occur because sperm are too efficient at reaching and fertilizing eggs due to the selective pressures of sperm competition. Such a situation is often deleterious to the female: in other words, the male–male competition among sperm spills over to create sexual conflict.

Sperm motility describes the ability of sperm to move properly through the female reproductive tract or through water to reach the egg. Sperm motility can also be thought of as the quality, which is a factor in successful conception; sperm that do not "swim" properly will not reach the egg in order to fertilize it. Sperm motility in mammals also facilitates the passage of the sperm through the cumulus oophorus and the zona pellucida, which surround the mammalian oocyte.
Sperm guidance is the process by which sperm cells (spermatozoa) are directed to the oocyte (egg) for the aim of fertilization. In the case of marine invertebrates the guidance is done by chemotaxis. In the case of mammals, it appears to be done by chemotaxis, thermotaxis and rheotaxis.
Sperm chemotaxis is a form of sperm guidance, in which sperm cells (spermatozoa) follow a concentration gradient of a chemoattractant secreted from the oocyte and thereby reach the oocyte.
Oocyteactivation is a series of processes that occur in the oocyte during fertilization.

Most mammals are viviparous, giving birth to live young. However, the five species of monotreme, the platypuses and the echidnas, lay eggs. The monotremes have a sex determination system different from that of most other mammals. In particular, the sex chromosomes of a platypus are more like those of a chicken than those of a therian mammal.
David B. Dusenbery is a biophysicist with a central interest in how information influences the behavior of organisms. In later years, he also considered the physical constraints hydrodynamics imposes on microorganisms and gametes.
A disintegrin and metalloprotease 3, or ADAM3, belongs to a family of peptidase proteins referred to as ADAMs. Many of these are solely found in spermatogenic cells, specifically in the anterior portion of capacitated spermatozoa heads. This membrane protein is critical for crucial steps in fertilization such as migration of sperm through the uterus to the oviduct as well as binding to the zona pellucida. Inactivation of ADAM3 is a cause of male infertility.

Protists are the eukaryotes that cannot be classified as plants, fungi or animals. They are mostly unicellular and microscopic. Many unicellular protists, particularly protozoans, are motile and can generate movement using flagella, cilia or pseudopods. Cells which use flagella for movement are usually referred to as flagellates, cells which use cilia are usually referred to as ciliates, and cells which use pseudopods are usually referred to as amoeba or amoeboids. Other protists are not motile, and consequently have no built-in movement mechanism.
Michael Eisenbach, Ph.D., is an Israeli biochemist who specializes in the navigation mechanisms of bacterial and sperm cells. He is a professor emeritus at the Weizmann Institute of Science, Department of Biomolecular Sciences, Rehovot, Israel. He discovered that sperm cells (spermatozoa) of mammals are actively guided to the egg. This opened the research field of mammalian sperm navigation. He demonstrated that the active navigation entails chemotaxis and thermotaxis. He made seminal contributions to the understanding of these two processes at the molecular, physiological and behavioural levels, as well as contributing to our understanding of the molecular mechanism of bacterial chemotaxis.