Related Research Articles

Ubiquitin is a small regulatory protein found in most tissues of eukaryotic organisms, i.e., it is found ubiquitously. It was discovered in 1975 by Gideon Goldstein and further characterized throughout the late 1970s and 1980s. Four genes in the human genome code for ubiquitin: UBB, UBC, UBA52 and RPS27A.
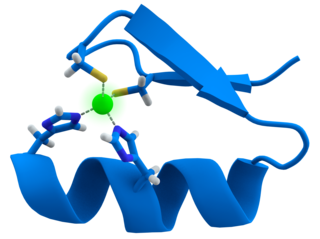
A zinc finger is a small protein structural motif that is characterized by the coordination of one or more zinc ions (Zn2+) which stabilizes the fold. It was originally coined to describe the finger-like appearance of a hypothesized structure from the African clawed frog (Xenopus laevis) transcription factor IIIA. However, it has been found to encompass a wide variety of differing protein structures in eukaryotic cells. Xenopus laevis TFIIIA was originally demonstrated to contain zinc and require the metal for function in 1983, the first such reported zinc requirement for a gene regulatory protein followed soon thereafter by the Krüppel factor in Drosophila. It often appears as a metal-binding domain in multi-domain proteins.

Histone acetyltransferases (HATs) are enzymes that acetylate conserved lysine amino acids on histone proteins by transferring an acetyl group from acetyl-CoA to form ε-N-acetyllysine. DNA is wrapped around histones, and, by transferring an acetyl group to the histones, genes can be turned on and off. In general, histone acetylation increases gene expression.

A ubiquitin ligase is a protein that recruits an E2 ubiquitin-conjugating enzyme that has been loaded with ubiquitin, recognizes a protein substrate, and assists or directly catalyzes the transfer of ubiquitin from the E2 to the protein substrate. In simple and more general terms, the ligase enables movement of ubiquitin from a ubiquitin carrier to another thing by some mechanism. The ubiquitin, once it reaches its destination, ends up being attached by an isopeptide bond to a lysine residue, which is part of the target protein. E3 ligases interact with both the target protein and the E2 enzyme, and so impart substrate specificity to the E2. Commonly, E3s polyubiquitinate their substrate with Lys48-linked chains of ubiquitin, targeting the substrate for destruction by the proteasome. However, many other types of linkages are possible and alter a protein's activity, interactions, or localization. Ubiquitination by E3 ligases regulates diverse areas such as cell trafficking, DNA repair, and signaling and is of profound importance in cell biology. E3 ligases are also key players in cell cycle control, mediating the degradation of cyclins, as well as cyclin dependent kinase inhibitor proteins. The human genome encodes over 600 putative E3 ligases, allowing for tremendous diversity in substrates.
SV40 large T antigen is a hexamer protein that is a dominant-acting oncoprotein derived from the polyomavirus SV40. TAg is capable of inducing malignant transformation of a variety of cell types. The transforming activity of TAg is due in large part to its perturbation of the retinoblastoma (pRb) and p53 tumor suppressor proteins. In addition, TAg binds to several other cellular factors, including the transcriptional co-activators p300 and CBP, which may contribute to its transformation function. Similar proteins from related viruses are known as large tumor antigen in general.

Mouse double minute 2 homolog (MDM2) also known as E3 ubiquitin-protein ligase Mdm2 is a protein that in humans is encoded by the MDM2 gene. Mdm2 is an important negative regulator of the p53 tumor suppressor. Mdm2 protein functions both as an E3 ubiquitin ligase that recognizes the N-terminal trans-activation domain (TAD) of the p53 tumor suppressor and as an inhibitor of p53 transcriptional activation.

Dystroglycan is a protein that in humans is encoded by the DAG1 gene.
Ubiquitin-conjugating enzymes, also known as E2 enzymes and more rarely as ubiquitin-carrier enzymes, perform the second step in the ubiquitination reaction that targets a protein for degradation via the proteasome. The ubiquitination process covalently attaches ubiquitin, a short protein of 76 amino acids, to a lysine residue on the target protein. Once a protein has been tagged with one ubiquitin molecule, additional rounds of ubiquitination form a polyubiquitin chain that is recognized by the proteasome's 19S regulatory particle, triggering the ATP-dependent unfolding of the target protein that allows passage into the proteasome's 20S core particle, where proteases degrade the target into short peptide fragments for recycling by the cell.

CREB-binding protein, also known as CREBBP or CBP or KAT3A, is a coactivator encoded by the CREBBP gene in humans, located on chromosome 16p13.3. CBP has intrinsic acetyltransferase functions; it is able to add acetyl groups to both transcription factors as well as histone lysines, the latter of which has been shown to alter chromatin structure making genes more accessible for transcription. This relatively unique acetyltransferase activity is also seen in another transcription enzyme, EP300 (p300). Together, they are known as the p300-CBP coactivator family and are known to associate with more than 16,000 genes in humans; however, while these proteins share many structural features, emerging evidence suggests that these two co-activators may promote transcription of genes with different biological functions.
Dystrobrevin is a protein that binds to dystrophin in the costamere of skeletal muscle cells. In humans, there are at least two isoforms of dystrobrevin, dystrobrevin alpha and dystrobrevin beta.

RING-box protein 2 is a protein that in humans is encoded by the RNF7 gene.

The human gene UBR1 encodes the enzyme ubiquitin-protein ligase E3 component n-recognin 1.

Tripartite motif-containing protein 32 is a protein that in humans is encoded by the TRIM32 gene. Since its discovery in 1995, TRIM32 has been shown to be implicated in a number of diverse biological pathways.

Tripartite motif-containing 22, also known as TRIM22, is a protein which in humans is encoded by the TRIM22 gene.

In molecular biology, a RING (short for Really Interesting New Gene) finger domain is a protein structural domain of zinc finger type which contains a C3HC4 amino acid motif which binds two zinc cations (seven cysteines and one histidine arranged non-consecutively). This protein domain contains 40 to 60 amino acids. Many proteins containing a RING finger play a key role in the ubiquitination pathway.

The BTB/POZ domain is a structural domain found in proteins across the domain Eukarya. Given its prevalence in eukaryotes and its absence in Archaea and bacteria, it likely arose after the origin of eukaryotes. While primarily a protein-protein interaction domain, some BTB domains have additional functionality in transcriptional regulation, cytoskeletal mobility, protein ubiquitination and degradation, and ion channel formation and operation. BTB domains have traditionally been classified by the other structural features present in the protein.

In molecular biology, TAZ zinc finger domains are zinc-containing domains found in the homologous transcriptional co-activators CREB-binding protein (CBP) and the P300. CBP and P300 are histone acetyltransferases that catalyse the reversible acetylation of all four histones in nucleosomes, acting to regulate transcription via chromatin remodelling. These large nuclear proteins interact with numerous transcription factors and viral oncoproteins, including p53 tumour suppressor protein, E1A oncoprotein, MyoD, and GATA-1, and are involved in cell growth, differentiation and apoptosis. Both CBP and P300 have two copies of the TAZ domain, one in the N-terminal region, the other in the C-terminal region. The TAZ1 domain of CBP and P300 forms a complex with CITED2, inhibiting the activity of the hypoxia inducible factor (HIF-1alpha) and thereby attenuating the cellular response to low tissue oxygen concentration. Adaptation to hypoxia is mediated by transactivation of hypoxia-responsive genes by hypoxia-inducible factor-1 (HIF-1) in complex with the CBP and p300 transcriptional coactivators.

In molecular biology the B-box-type zinc finger domain is a short protein domain of around 40 amino acid residues in length. B-box zinc fingers can be divided into two groups, where types 1 and 2 B-box domains differ in their consensus sequence and in the spacing of the 7-8 zinc-binding residues. Several proteins contain both types 1 and 2 B-boxes, suggesting some level of cooperativity between these two domains.

In molecular biology, the Btk-type zinc finger or Btk motif (BM) is a conserved zinc-binding motif containing conserved cysteines and a histidine that is present in certain eukaryotic signalling proteins. The motif is named after Bruton's tyrosine kinase (Btk), an enzyme which is essential for B cell maturation in humans and mice. Btk is a member of the Tec family of protein tyrosine kinases (PTK). These kinases contain a conserved Tec homology (TH) domain between the N-terminal pleckstrin homology (PH) domain and the Src homology 3 (SH3) domain. The N-terminal of the TH domain is highly conserved and known as the Btf motif, while the C-terminal region of the TH domain contains a proline-rich region (PRR). The Btk motif contains a conserved His and three Cys residues that form a zinc finger, while PRRs are commonly involved in protein-protein interactions, including interactions with G proteins. The TH domain may be of functional importance in various signalling pathways in different species. A complete TH domain, containing both the Btk and PRR regions, has not been found outside the Tec family; however, the Btk motif on its own does occur in other proteins, usually C-terminal to a PH domain.

Ubiquitin-binding domains (UBDs) are protein domains that recognise and bind non-covalently to ubiquitin through protein-protein interactions. As of 2019, a total of 29 types of UBDs had been identified in the human proteome. Most UBDs bind to ubiquitin only weakly, with binding affinities in the low to mid μM range. Proteins containing UBDs are known as ubiquitin-binding proteins or sometimes as "ubiquitin receptors".
References
- ↑ Ponting CP, Blake DJ, Davies KE, Kendrick-Jones J, Winder SJ (January 1996). "ZZ and TAZ: new putative zinc fingers in dystrophin and other proteins". Trends Biochem. Sci. 21 (1): 11–13. doi:10.1016/s0968-0004(06)80020-4. PMID 8848831.
- 1 2 Legge GB, Martinez-Yamout MA, Hambly DM, Trinh T, Lee BM, Dyson HJ, Wright PE (October 2004). "ZZ domain of CBP: an unusual zinc finger fold in a protein interaction module". J. Mol. Biol. 343 (4): 1081–93. doi:10.1016/j.jmb.2004.08.087. PMID 15476823.
- ↑ Kang X, Chong J, Ni M (March 2005). "HYPERSENSITIVE TO RED AND BLUE 1, a ZZ-type zinc finger protein, regulates phytochrome B-mediated red and cryptochrome-mediated blue light responses". Plant Cell. 17 (3): 822–35. doi:10.1105/tpc.104.029165. PMC 1069701 . PMID 15705950.
- ↑ Nishito Y, Hasegawa M, Inohara N, Núñez G (June 2006). "MEX is a testis-specific E3 ubiquitin ligase that promotes death receptor-induced apoptosis". Biochem. J. 396 (3): 411–7. doi:10.1042/BJ20051814. PMC 1482824 . PMID 16522193.
- ↑ Hnia K, Zouiten D, Cantel S, Chazalette D, Hugon G, Fehrentz JA, Masmoudi A, Diment A, Bramham J, Mornet D, Winder SJ (February 2007). "ZZ domain of dystrophin and utrophin: topology and mapping of a beta-dystroglycan interaction site". Biochem. J. 401 (3): 667–77. doi:10.1042/BJ20061051. PMC 1770854 . PMID 17009962.