
Haplogroup X is a human mitochondrial DNA (mtDNA) haplogroup. It is found in North America, Europe, Western Asia, North Africa, and the Horn of Africa.
Haplogroup K, formerly Haplogroup UK, is a human mitochondrial DNA (mtDNA) haplogroup. It is defined by the HVR1 mutations 16224C and 16311C. It is now known that K is a subclade of U8.
Haplogroup T is a human mitochondrial DNA (mtDNA) haplogroup. It is believed to have originated around 25,100 years ago in the Near East.
Haplogroup V is a human mitochondrial DNA (mtDNA) haplogroup. The clade is believed to have originated over 14,000 years ago in Southern Europe.
Haplogroup HV is a human mitochondrial DNA (mtDNA) haplogroup.
Haplogroup U is a human mitochondrial DNA haplogroup (mtDNA). The clade arose from haplogroup R, likely during the early Upper Paleolithic. Its various subclades are found widely distributed across Northern and Eastern Europe, Central, Western and South Asia, as well as North Africa, the Horn of Africa, and the Canary Islands.

Haplogroup N is a human mitochondrial DNA (mtDNA) clade. A macrohaplogroup, its descendant lineages are distributed across many continents. Like its sibling macrohaplogroup M, macrohaplogroup N is a descendant of the haplogroup L3.
In human mitochondrial genetics, Haplogroup C is a human mitochondrial DNA (mtDNA) haplogroup.
Haplogroup L2 is a human mitochondrial DNA (mtDNA) haplogroup with a widespread modern distribution, particularly in Subequatorial Africa. Its L2a subclade is a somewhat frequent and widely distributed mtDNA cluster on the continent, as well as among those in the Americas.
Haplogroup I is a human mitochondrial DNA (mtDNA) haplogroup. It is believed to have originated about 21,000 years ago, during the Last Glacial Maximum (LGM) period in West Asia. The haplogroup is unusual in that it is now widely distributed geographically, but is common in only a few small areas of East Africa, West Asia and Europe. It is especially common among the El Molo and Rendille peoples of Kenya, various regions of Iran, the Lemko people of Slovakia, Poland and Ukraine, the island of Krk in Croatia, the department of Finistère in France and some parts of Scotland and Ireland.

Haplogroup N (M231) is a Y-chromosome DNA haplogroup defined by the presence of the single-nucleotide polymorphism (SNP) marker M231.
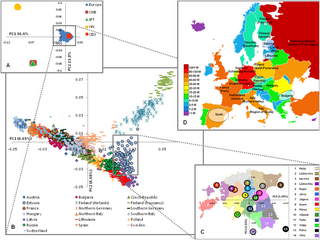
The genetic history of Europe includes information around the formation, ethnogenesis, and other DNA-specific information about populations indigenous, or living in Europe.
In human mitochondrial genetics, Haplogroup G is a human mitochondrial DNA (mtDNA) haplogroup.
Haplogroup H is a human mitochondrial DNA (mtDNA) haplogroup. The clade is believed to have originated in Southwest Asia, near present day Syria, around 20,000 to 25,000 years ago. Mitochondrial haplogroup H is today predominantly found in Europe, and is believed to have evolved before the Last Glacial Maximum (LGM). It first expanded in the northern Near East and Southern Caucasus soon, and later migrations from Iberia suggest that the clade reached Europe before the Last Glacial Maximum. The haplogroup has also spread to parts of Africa, Siberia and Inner Asia. Today, around 40% of all maternal lineages in Europe belong to haplogroup H.

Haplogroup R1a, or haplogroup R-M420, is a human Y-chromosome DNA haplogroup which is distributed in a large region in Eurasia, extending from Scandinavia and Central Europe to Central Asia, southern Siberia and South Asia.

Haplogroup R1b (R-M343), previously known as Hg1 and Eu18, is a human Y-chromosome haplogroup.

African admixture in Europe refers to the presence of human genotypes attributable to periods of human population dispersals out of Africa in the genetic history of Europe. For example, certain Y-DNA and mtDNA lineages are thought to have spread from Northeastern Africa to the Near East during the later Pleistocene, and from there to Europe with the Neolithic Revolution.
Population genetics is a scientific discipline which contributes to the examination of the human evolutionary and historical migrations. Particularly useful information is provided by the research of two uniparental markers within our genome, the Y-chromosome (Y-DNA) and mitochondrial DNA (mtDNA), as well as autosomal DNA. The data from Y-DNA and autosomal DNA suggests that the Croats mostly are descendants of the Slavs of the medieval migration period, according to mtDNA have genetic diversity which fits within a broader European maternal genetic landscape, and overall have a uniformity with other South Slavs from the territory of former Yugoslavia.

Haplogroup R-M269 is the sub-clade of human Y-chromosome haplogroup R1b that is defined by the SNP marker M269. According to ISOGG 2020 it is phylogenetically classified as R1b1a1b. It underwent intensive research and was previously classified as R1b1a2, R1b1c, R1b1b2 and R1b1a1a2.
Early European Farmers (EEF), First European Farmers, Neolithic European Farmers, Ancient Aegean Farmers, or Anatolian Neolithic Farmers are names used to describe a distinct group of early Neolithic farmers who brought agriculture to Europe and Northwest Africa (Maghreb). Although the spread of agriculture from the Middle East to Europe has long been recognised through archaeology, it is only recent advances in archaeogenetics that have confirmed that this spread was strongly correlated with a migration of these farmers, and was not just a cultural exchange.
