
Branchiopoda is a class of crustaceans. It comprises fairy shrimp, clam shrimp, Diplostraca, Notostraca, the Devonian Lepidocaris and possibly the Cambrian Rehbachiella. They are mostly small, freshwater animals that feed on plankton and detritus.

Amphipoda is an order of malacostracan crustaceans with no carapace and generally with laterally compressed bodies. Amphipods range in size from 1 to 340 millimetres and are mostly detritivores or scavengers. There are more than 9,900 amphipod species so far described. They are mostly marine animals, but are found in almost all aquatic environments. Some 1,900 species live in fresh water, and the order also includes the terrestrial sandhoppers such as Talitrus saltator and Arcitalitrus sylvaticus.

Isopoda is an order of crustacean, which includes woodlice and their relatives. Members of this group are called Isopods and include both terrestrial and aquatic species. All have rigid, segmented exoskeletons, two pairs of antennae, seven pairs of jointed limbs on the thorax, and five pairs of branching appendages on the abdomen that are used in respiration. Females brood their young in a pouch under their thorax.

The family Argulidae, whose members are commonly known as carp lice or fish lice, are parasitic crustaceans in the class Ichthyostraca. It is the only family in the monotypic subclass Branchiura and the order Arguloida, although a second family, Dipteropeltidae, has been proposed. Although they are thought to be primitive forms, they have no fossil record.
Traditionally, the Myodocopa and Podocopa have been classified as subclasses within the class Ostracoda, although there is some question about how closely related the two groups actually are. The Myodocopa are defined by possession of a poorly calcified carapace, and 8–9 articles in the exopod of the second antenna. The ventral margin of the carapace is not concave, and the valves do not overlap to a great extent.
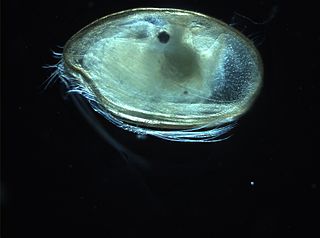
The Myodocopida is one of the two orders within the Myodocopa, in turn a subclass of the Ostracoda. The Myodocopida are distinguished by a worm-like seventh limb, and, usually, a rostrum above an incisure (notch) from which the antennae can protrude. Unlike other ostracods, many species of the Myodocopida have lateral compound eyes Over the last thirty years there has been much research into the morphology, behaviour and distribution of myodocopids. More recently, DNA sequences have been used to investigate the phylogeny of various groups.

Cylindroleberididae is a family of ostracods that shows remarkable morphological diversity. The defining feature is the possession of gills: 7–8 leaf-like pairs at the posterior of the body. Other features common to all species in the family include a "baleen-comb" on both the maxilla and the fifth limb, a sword-shaped coxal endite on the mandible, and the triaenid bristles on the basal endites of the mandible.
The Halocyprida is one of the two orders within the Myodocopa, in turn a subclass of the ostracods. Like their relatives in the order Myodocopida, they have a long exopod on the second antenna. However, unlike myodocopids, their fifth appendage is leg-like rather than modified for feeding, their seventh limb is reduced or absent, and they have no lateral eyes. The group is primarily planktonic. There are two suborders: Halocypridina and Cladocopina.

The Podocopa are a subclass of ostracods. Members of the subclass Podocopa can be differentiated from the other subclass of ostracods (Myodocopa) by the morphology of the second antenna: the Podocopa have a relatively long endopod, whereas the Myodocopa have a relatively long exopod. The seventh limb of the Podocopa has a variety of forms or is absent, but is never an annulated worm-like limb.

The Podocopida are an order of ostracods in the subclass Podocopa. It is the most diverse of the five orders of ostracods, and the only one with freshwater species. The group also has a rich fossil record.

Gigantocypris, sometimes known as giant ostracod or giant seed shrimp, is a genus of ostracod crustaceans in family Cypridinidae, and among the most well-known members of the class Ostracoda. Its members are extremely large for ostracods, measuring up to 3.2 cm (1.3 in) across, have a globular shape, are typically semi-transparent orange or reddish, and have a large pair of mirror-like eyes that are used to locate their small animal prey. They are found worldwide in dark, deep and cold oceans.

Vargula hilgendorfii, sometimes called the sea-firefly and one of three bioluminescent species known in Japan as umi-hotaru (海蛍), is a species of ostracod crustacean. It is the only member of genus Vargula to inhabit Japanese waters; all other members of its genus inhabit the Gulf of Mexico, the Caribbean Sea, and waters off the coast of California. V. hilgendorfii was formerly more common, but its numbers have fallen significantly.
Potamocypris unicaudata is a species of ostracod crustacean in the family Cyprididae, subfamily Cypridopsinae. It is abundantly found in ditches and ponds near the sea shore, where freshwater slightly mingles with sea water. It is known from both Europe and North America.
Potamocypris smaragdina is a species of ostracod crustacean in the family Cyprididae, subfamily Cypridopsinae. It is known from both Europe and North America.

Crustaceans are a group of arthropods that are a part of the subphylum Crustacea, a large, diverse group of mainly aquatic arthropods including decapods, seed shrimp, branchiopods, fish lice, krill, remipedes, isopods, barnacles, copepods, opossum shrimps, amphipods and mantis shrimp. The crustacean group can be treated as a subphylum under the clade Mandibulata. It is now well accepted that the hexapods emerged deep in the Crustacean group, with the completed group referred to as Pancrustacea. The three classes Cephalocarida, Branchiopoda and Remipedia are more closely related to the hexapods than they are to any of the other crustaceans.
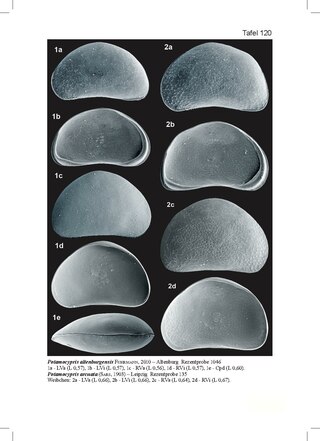
Potamocypris arcuata is a species of ostracod crustacean in the family Cyprididae, subfamily Cypridopsinae. It is mainly known from the southern areas of the Palaearctic.
The biological systematics and taxonomy of invertebrates as proposed by Richard C. Brusca and Gary J. Brusca in 2003 is a system of classification of invertebrates, as a way to classify animals without backbones.
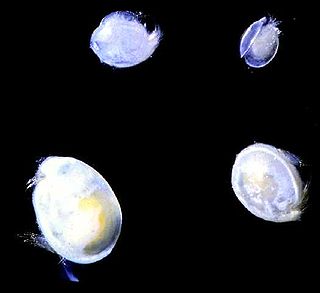
Oligostraca is a superclass of crustaceans. It consist of the following three classes:
Enewton is a genus in the family Cypridinidae. The genus contains bioluminescent species, and in particular is one of the Caribbean genera of bioluminescent ostracods that perform stereotyped bioluminescent mating displays.





