p14ARF (also called ARF tumor suppressor, ARF, p14ARF) is an alternate reading frame protein product of the CDKN2A locus (i.e. INK4a/ARF locus).[1] p14ARF is induced in response to elevated mitogenic stimulation, such as aberrant growth signaling from MYC and Ras (protein).[2] It accumulates mainly in the nucleolus where it forms stable complexes with NPM or Mdm2. These interactions allow p14ARF to act as a tumor suppressor by inhibiting ribosome biogenesis or initiating p53-dependent cell cycle arrest and apoptosis, respectively.[3] p14ARF is an atypical protein, in terms of its transcription, its amino acid composition, and its degradation: it is transcribed in an alternate reading frame of a different protein, it is highly basic,[1] and it is polyubiquinated at the N-terminus.[4]
Both p16INK4a and p14ARF are involved in cell cycle regulation. p14ARF inhibits mdm2, thus promoting p53, which promotes p21 activation, which then binds and inactivates certain cyclin-CDK complexes, which would otherwise promote transcription of genes that would carry the cell through the G1/S checkpoint of the cell cycle. Loss of p14ARF by a homozygous mutation in the CDKN2A (INK4A) gene will lead to elevated levels in mdm2 and, therefore, loss of p53 function and cell cycle control.
The equivalent in mice is p19ARF.
Background
The p14ARF transcript was first identified in humans in 1995,[5][6] and its protein product confirmed in mice that same year.[7] Its gene locus is on the short arm of chromosome 9 in humans, and on a corresponding location on chromosome 4 in mice.[1] It is located near the genes for the tandem repeats INK4a and INK4b, which are 16 kDa (p16INK4a) and 15 kDa (p15INK4b) proteins, respectively. These INK4 proteins directly inhibit the cyclin D-dependent kinases CDK4 and CDK6. There are other INK4 genes on other chromosomes, however these are not linked to cancer, and so their functions are not likely to be overlapping. An important cyclin-dependent substrate is the retinoblastoma protein Rb, which is phosphorylated in late gap 1 phase (G1 phase), allowing G1 exit. The Rb protein limits cell proliferation by blocking the activity of E2F transcription factors, which activate the transcription of genes needed for DNA replication. When Rb is phosphorylated by cyclin D and E-dependent kinases during the G1 phase of the cell cycle, Rb can not block E2F-dependent transcription, and the cell can progress to the DNA synthetic phase(S phase).[8] Therefore, INK4a and INK4b serve as tumor suppressors by restricting proliferation though the inhibition of the CDKs responsible for Rb phosphorylation.[7]
In addition to the INK4a protein, the unrelated protein, ARF, is transcribed from an alternate reading frame at the INK4a/ARF locus.[1] INK4a and p14ARF mRNA each consist of three exons. They share exons 2 and 3, but there are two different exon 1 transcripts, α and β. Exon 1β (E1β) is intercalated between the genes for INK4a and INK4b.[1] Although exon 1α (E1α) and E1β are about the same in terms of content and size, the 5’ AUG (start codon) of exon 1β has its own promoter and opens an alternative reading frame in exon 2, hence the name p14ARF (ARF exon 3 is not translated). Because of this, INK4a and p14ARF have unrelated amino acid sequences despite overlapping coding regions and have distinct functions. This dual-use of coding sequences is not commonly seen in mammals, making p14ARF an unusual protein.[1] When the ARF β-transcript was found, it was thought that it probably would not encode a protein.[5][6] In humans, ARF is translated into the 14kDa, 132 amino acid [[p14ARF]] protein, and in mice, it is translated into the 19kDa, 169 amino acid p19Arf.[1] The E1β protein segment of mouse and human ARF are 45% identical, with an overall ARF identity of 50%, compared to a 72% identity between mouse and human INK4a E1α segment, and a 65% overall identity.[7]
Although the INK4a and ARF proteins are structurally and functionally different, they are both involved in cell cycle progression. Together, their broad inhibitory role may help counter oncogenic signals. As mentioned above, INK4a inhibits proliferation by indirectly allowing Rb to remain associated with E2F transcription factors. ARF is involved in p53 activation by inhibiting Mdm2 (HDM2 in humans).[8] Mdm2 binds to p53, inhibiting its transcriptional activity. Mdm2 also has E3 ubiquitin ligase activity toward p53, and promotes its exportation from the cell nucleus to the cytoplasm for degradation. By antagonizing Mdm2, ARF permits the transcriptional activity of p53 that would lead to cell cycle arrest or apoptosis. A loss of ARF or p53, therefore, would give cells a survival advantage.[1]
The function of ARF has primarily been attributed to its Mdm2/p53 mechanism. ARF does, however, also inhibit proliferation in cells lacking p53 or p53 and Mdm2.[9] In 2004 has been found that one of ARF's p53-independent functions involves its binding to nucleophosmin/B23 (NPM).[9] NPM is an acidic ribosomalchaperone (protein) involved in preribosomal processing and nuclear exportation independent of p53, and oligomerizes with itself and p14ARF. Nearly half of p14ARF is found in NPM-containing complexes with high molecular mass (2 to 5 MDa). Enforced expression of ARF retards early 47S/45S rRNA precursor processing and inhibits 32S rRNA cleavage. This suggests that p14ARF can bind to NPM, inhibiting rRNA processing.[9] ARF-null cells have increased nucleolar area, increased ribosome biogenesis, and a corresponding increase in protein synthesis.[10] The larger size resulting from more ribosomes and protein is not associated with increased proliferation, however, and this ARF-null phenotype occurs even though the normal basal levels of Arf are usually low. Knocking down ARF with siRNA to exon 1β results in increased rRNA transcripts, rRNA processing, and ribosome nuclear export. The unrestrained ribosome biogenesis seen when NPM is not bound to ARF does not occur if NPM is also absent. Although the induction of ARF in response to oncogenic signals is considered to be of primary importance, the low levels of ARF seen in interphase cells also has a considerable effect in terms of keeping cell growth in check. Therefore, the function of basal level ARF in the NPM/ARF complex appears to be to monitor steady-state ribosome biogenesis and growth independently of preventing proliferation.[10]
Role in Disease
Very commonly, cancer is associated with a loss of function of INK4a, ARF, Rb, or p53.[11] Without INK4a, Cdk4/6 can inappropriately phosphorylate Rb, leading to increased E2F-dependent transcription. Without ARF, Mdm2 can inappropriately inhibit p53, leading to increased cell survival.
The INK4a/ARF locus is found to be deleted or silenced in many kinds of tumors. For example, of the 100 primary breast carcinomas, approximately 41% have p14ARF defects.[12] In a separate study, 32% of colorectal adenomas (non-cancerous tumors) were found to have p14ARF inactivation due to hypermethylation of the promoter. Mouse models lacking p19Arf, p53, and Mdm2 are more prone to tumor development than mice without Mdm2 and p53, alone. This suggests that p19Arf has Mdm2- and p53-independent effects, as well.[13] Investigating this idea lead to the recent discovery of smARF.[14]
Homozygous deletions and other mutations of CDK2NA (ARF) have been found to be associated with glioblastoma.[15]
smARF
Until recently, the two known effects of ARF were growth inhibition by NPM interactions and apoptosis induction by Mdm2 interactions. The function of ARF involving p53-independent death, has now been attributed to the small mitochondrialisoform of ARF, smARF.[14] While full-length ARF inhibits cell growth by cell cycle arrest or type I apoptotic death, smARF kills cells by type II autophagic death. Like ARF, the expression of smARF increases when there are aberrant proliferation signals. When smARF is overexpressed, it localizes to the mitochondrial matrix, damaging the mitochondria membrane potential and structure, and leading to autophagic cell death.[16]
The translation of the truncated ARF, smARF, is initiated at an internal methionine (M45) of the ARF transcript in human and mouse cells. SmARF is also detected in rats, even though an internal methionine is not present in the rat transcript. This suggests that there is an alternate mechanism to form smARF, underscoring the importance of this isoform.[14] The role of smARF is distinct from that of ARF, as it lacks the nuclear localization signal (NLS) and cannot bind to Mdm2 or NPM.[3] In some cell types, however, full-length ARF can also localize to the mitochondria and induce type II cell death, suggesting that in addition to autophagy being a starvation or other environmental response, it may also be involved in responding to oncogene activation.[2]
Biochemistry
ARF expression is regulated by oncogenic signaling. Aberrant mitogenic stimulation, such as by MYC or Ras (protein), will increase its expression, as will an amplification of mutated p53 or Mdm2, or p53 loss.[8] ARF can also be induced by enforced E2F expression. Although E2F expression is increased during the cell cycle, ARF expression probably is not because the activation of a second, unknown transcription factor might be needed to prevent an ARF response to transient E2F increases.[11] ARF is negatively regulated by Rb-E2F complexes [11] and by amplified p53 activation.[8] Aberrant growth signals also increase smARF expression.[16]
ARF is a highly basic (pI>12) and hydrophobic protein.[8] Its basic nature is attributed to its arginine content; more than 20% of its amino acids are arginine, and it contains little or no lysine. Due to these characteristics, ARF is likely to be unstructured unless it is bound to other targets. It reportedly complexes with more than 25 proteins, although the significance of each of these interactions is not known.[1] One of these interactions results in sumoylating activity, suggesting that ARF may modify proteins to which it binds. The SUMO protein is a small ubiquitin-like modifier, which is added to lysly ε-amino groups. This process involves a three-enzyme cascade similar to the way ubiquitylation occurs. E1 is an activating enzyme, E2 is a conjugation enzyme, and E3 is a ligase. ARF associates with UBC9, the only SUMO E2 known, suggesting ARF facilitates SUMO conjugation. The importance of this role is unknown, as sumoylation is involved in different functions, such as protein trafficking, ubiquitylation interference, and gene expression changes.[1]
The half-life of ARF is about 6 hours,[4] while the half-life of smARF is less than 1 hour.[3] Both isoforms are degraded in the proteasome.[1][4] ARF is targeted for the proteasome by N-terminusubiquitylation.[4] Proteins are usually ubiquinated at lysine residues. Human [[p14ARF]], however, does not contain any lysines, and mouse p19Arf only contains one lysine. If the mouse lysine is replaced with arginine, there is no effect on its degradation, suggesting it is also ubiquinated at the N-terminus. This adds to the uniqueness of the ARF proteins, because most eukaryotic proteins are acetylated at the N-terminus, preventing ubiquination at this location. Penultimate residues affect the efficiency of acetylation, in that acetylation is promoted by acidic residues and inhibited by basic ones. The N-terminal amino acid sequences of p19Arf (Met-Gly-Arg) and p14ARF (Met-Val-Arg) would be processed by methionine aminopeptidase but would not be acetylated, allowing ubiquination to proceed. The sequence of smARF, however, predicts that the initiating methionine would not be cleaved by methionine aminopeptidase and would probably be acetylated, and so is degraded by the proteasome without ubiquination.[1]
Full-length nucleolar ARF appears to be stabilized by NPM. The NPM-ARF complex does not block the N-terminus of ARF but likely protects ARF from being accessed by degradation machinery.[4] The mitochondrial matrix protein p32 stabilizes smARF.[16] This protein binds various cellular and viral proteins, but its exact function is unknown. Knocking down p32 dramatically decreases smARF levels by increasing its turnover. The levels of p19Arf are not affected by p32 knockdown, and so p32 specifically stabilizes smARF, possibly by protecting it from the proteasome or from mitochondrialproteases.[16]
Related Research Articles
The cell cycle, or cell-division cycle, is the series of events that take place in a cell that cause it to divide into two daughter cells. These events include the duplication of its DNA and some of its organelles, and subsequently the partitioning of its cytoplasm and other components into two daughter cells in a process called cell division.
Tumor protein P53, also known as p53, cellular tumor antigen p53, the Guardian of the Genome, phosphoprotein p53, tumor suppressor p53, antigen NY-CO-13, or transformation-related protein 53 (TRP53), is any isoform of a protein encoded by homologous genes in various organisms, such as TP53 (humans) and Trp53 (mice). The p53 proteins are crucial in vertebrates, where they prevent cancer formation. As such, p53 has been described as "the guardian of the genome" because of its role in conserving stability by preventing genome mutation. Hence TP53 is classified as a tumor suppressor gene.
A tumor suppressor gene (TSG), or anti-oncogene, is a gene that regulates a cell during cell division and replication. If the cell grows uncontrollably, it will result in cancer. When a tumor suppressor gene is mutated, it results in a loss or reduction in its function. In combination with other genetic mutations, this could allow the cell to grow abnormally. The loss of function for these genes may be even more significant in the development of human cancers, compared to the activation of oncogenes.
p73 is a protein related to the p53 tumor protein. Because of its structural resemblance to p53, it has also been considered a tumor suppressor. It is involved in cell cycle regulation, and induction of apoptosis. Like p53, p73 is characterized by the presence of different isoforms of the protein. This is explained by splice variants, and an alternative promoter in the DNA sequence.
Mouse double minute 2 homolog (MDM2) also known as E3 ubiquitin-protein ligase Mdm2 is a protein that in humans is encoded by the MDM2 gene. Mdm2 is an important negative regulator of the p53 tumor suppressor. Mdm2 protein functions both as an E3 ubiquitin ligase that recognizes the N-terminal trans-activation domain (TAD) of the p53 tumor suppressor and as an inhibitor of p53 transcriptional activation.
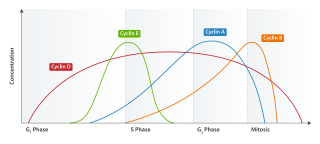
Cyclin E is a member of the cyclin family.
p16, is a protein that slows cell division by slowing the progression of the cell cycle from the G1 phase to the S phase, thereby acting as a tumor suppressor. It is encoded by the CDKN2A gene. A deletion in this gene can result in insufficient or non-functional p16, accelerating the cell cycle and resulting in many types of cancer.
INK4 is a family of cyclin-dependent kinase inhibitors (CKIs). The members of this family (p16INK4a, p15INK4b, p18INK4c, p19INK4d) are inhibitors of CDK4 (hence their name INhibitors of CDK4), and of CDK6. The other family of CKIs, CIP/KIP proteins are capable of inhibiting all CDKs. Enforced expression of INK4 proteins can lead to G1 arrest by promoting redistribution of Cip/Kip proteins and blocking cyclin E-CDK2 activity. In cycling cells, there is a resassortment of Cip/Kip proteins between CDK4/5 and CDK2 as cells progress through G1. Their function, inhibiting CDK4/6, is to block progression of the cell cycle beyond the G1 restriction point. In addition, INK4 proteins play roles in cellular senescence, apoptosis and DNA repair.
Cyclin D is a member of the cyclin protein family that is involved in regulating cell cycle progression. The synthesis of cyclin D is initiated during G1 and drives the G1/S phase transition. Cyclin D protein is anywhere from 155 to 477 amino acids in length.
Cyclin-dependent kinase 4 also known as cell division protein kinase 4 is an enzyme that in humans is encoded by the CDK4 gene. CDK4 is a member of the cyclin-dependent kinase family.
Cell division protein kinase 6 (CDK6) is an enzyme encoded by the CDK6 gene. It is regulated by cyclins, more specifically by Cyclin D proteins and Cyclin-dependent kinase inhibitor proteins. The protein encoded by this gene is a member of the cyclin-dependent kinase, (CDK) family, which includes CDK4. CDK family members are highly similar to the gene products of Saccharomyces cerevisiae cdc28, and Schizosaccharomyces pombe cdc2, and are known to be important regulators of cell cycle progression in the point of regulation named R or restriction point.
Acute myeloblastic leukemia with maturation (M2) is a subtype of acute myeloid leukemia (AML).
Nucleophosmin (NPM), also known as nucleolar phosphoprotein B23 or numatrin, is a protein that in humans is encoded by the NPM1 gene.
G1/S-specific cyclin-D2 is a protein that in humans is encoded by the CCND2 gene.
Cyclin-dependent kinase 4 inhibitor B also known as multiple tumor suppressor 2 (MTS-2) or p15INK4b is a protein that is encoded by the CDKN2B gene in humans.
Cyclin-dependent kinase 4 inhibitor C is an enzyme that in humans is encoded by the CDKN2C gene.
Cyclin-dependent kinase 4 inhibitor D is an enzyme that in humans is encoded by the CDKN2D gene.
CDKN2A, also known as cyclin-dependent kinase inhibitor 2A, is a gene which in humans is located at chromosome 9, band p21.3. It is ubiquitously expressed in many tissues and cell types. The gene codes for two proteins, including the INK4 family member p16 and p14arf. Both act as tumor suppressors by regulating the cell cycle. p16 inhibits cyclin dependent kinases 4 and 6 and thereby activates the retinoblastoma (Rb) family of proteins, which block traversal from G1 to S-phase. p14ARF activates the p53 tumor suppressor. Somatic mutations of CDKN2A are common in the majority of human cancers, with estimates that CDKN2A is the second most commonly inactivated gene in cancer after p53. Germline mutations of CDKN2A are associated with familial melanoma, glioblastoma and pancreatic cancer. The CDKN2A gene also contains one of 27 SNPs associated with increased risk of coronary artery disease.
The retinoblastoma protein is a tumor suppressor protein that is dysfunctional in several major cancers. One function of pRb is to prevent excessive cell growth by inhibiting cell cycle progression until a cell is ready to divide. When the cell is ready to divide, pRb is phosphorylated, inactivating it, and the cell cycle is allowed to progress. It is also a recruiter of several chromatin remodeling enzymes such as methylases and acetylases.
The CIP/KIP family is one of two families of mammalian cyclin dependent kinase (CDK) inhibitors (CKIs) involved in regulating the cell cycle. The CIP/KIP family is made up of three proteins: p21cip1/waf1, P27kip1, p57kip2 These proteins share sequence homology at the N-terminal domain which allows them to bind to both the cyclin and CDK. Their activity primarily involves the binding and inhibition of G1/S- and S-Cdks; however, they have also been shown to play an important role in activating the G1-CDKs CDK4 and CDK6. In addition, more recent work has shown that CIP/KIP family members have a number of CDK-independent roles involving regulation of transcription, apoptosis, and the cytoskeleton.
Zhang, Y., Y. Xiong, and W.G. Yarbrough. ARF Promotes MDM2 Degradation and Stabilizes p53: ARF-INK4a Locus Deletion Impairs Both the Rb and p53 Tumor Suppression Pathways. Cell 1998, 92(6):725-34.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.