Related Research Articles

The cell cycle, or cell-division cycle, is the series of events that take place in a cell that causes it to divide into two daughter cells. These events include the duplication of its DNA and some of its organelles, and subsequently the partitioning of its cytoplasm, chromosomes and other components into two daughter cells in a process called cell division.

Telophase is the final stage in both meiosis and mitosis in a eukaryotic cell. During telophase, the effects of prophase and prometaphase are reversed. As chromosomes reach the cell poles, a nuclear envelope is re-assembled around each set of chromatids, the nucleoli reappear, and chromosomes begin to decondense back into the expanded chromatin that is present during interphase. The mitotic spindle is disassembled and remaining spindle microtubules are depolymerized. Telophase accounts for approximately 2% of the cell cycle's duration.
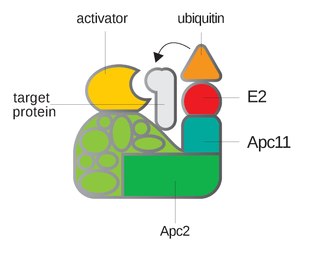
Anaphase-promoting complex is an E3 ubiquitin ligase that marks target cell cycle proteins for degradation by the 26S proteasome. The APC/C is a large complex of 11–13 subunit proteins, including a cullin (Apc2) and RING (Apc11) subunit much like SCF. Other parts of the APC/C have unknown functions but are highly conserved.
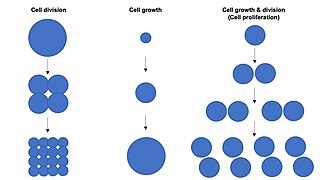
Cell growth refers to an increase in the total mass of a cell, including both cytoplasmic, nuclear and organelle volume. Cell growth occurs when the overall rate of cellular biosynthesis is greater than the overall rate of cellular degradation.

Cyclin-dependent kinases (CDKs) are the families of protein kinases first discovered for their role in regulating the cell cycle. They are also involved in regulating transcription, mRNA processing, and the differentiation of nerve cells. They are present in all known eukaryotes, and their regulatory function in the cell cycle has been evolutionarily conserved. In fact, yeast cells can proliferate normally when their CDK gene has been replaced with the homologous human gene. CDKs are relatively small proteins, with molecular weights ranging from 34 to 40 kDa, and contain little more than the kinase domain. By definition, a CDK binds a regulatory protein called a cyclin. Without cyclin, CDK has little kinase activity; only the cyclin-CDK complex is an active kinase but its activity can be typically further modulated by phosphorylation and other binding proteins, like p27. CDKs phosphorylate their substrates on serines and threonines, so they are serine-threonine kinases. The consensus sequence for the phosphorylation site in the amino acid sequence of a CDK substrate is [S/T*]PX[K/R], where S/T* is the phosphorylated serine or threonine, P is proline, X is any amino acid, K is lysine, and R is arginine.
Maturation-promoting factor (abbreviated MPF, also called mitosis-promoting factor or M-Phase-promoting factor) is the cyclin-Cdk complex that was discovered first in frog eggs. It stimulates the mitotic and meiotic phases of the cell cycle. MPF promotes the entrance into mitosis (the M phase) from the G2 phase by phosphorylating multiple proteins needed during mitosis. MPF is activated at the end of G2 by a phosphatase, which removes an inhibitory phosphate group added earlier.

G2 phase, Gap 2 phase, or Growth 2 phase, is the third subphase of interphase in the cell cycle directly preceding mitosis. It follows the successful completion of S phase, during which the cell’s DNA is replicated. G2 phase ends with the onset of prophase, the first phase of mitosis in which the cell’s chromatin condenses into chromosomes.
Endoreduplication is replication of the nuclear genome in the absence of mitosis, which leads to elevated nuclear gene content and polyploidy. Endoreplication can be understood simply as a variant form of the mitotic cell cycle (G1-S-G2-M) in which mitosis is circumvented entirely, due to modulation of cyclin-dependent kinase (CDK) activity. Examples of endoreplication characterized in arthropod, mammalian, and plant species suggest that it is a universal developmental mechanism responsible for the differentiation and morphogenesis of cell types that fulfill an array of biological functions. While endoreplication is often limited to specific cell types in animals, it is considerably more widespread in plants, such that polyploidy can be detected in the majority of plant tissues.

Cell cycle checkpoints are control mechanisms in the eukaryotic cell cycle which ensure its proper progression. Each checkpoint serves as a potential termination point along the cell cycle, during which the conditions of the cell are assessed, with progression through the various phases of the cell cycle occurring only when favorable conditions are met. There are many checkpoints in the cell cycle, but the three major ones are: the G1 checkpoint, also known as the Start or restriction checkpoint or Major Checkpoint; the G2/M checkpoint; and the metaphase-to-anaphase transition, also known as the spindle checkpoint. Progression through these checkpoints is largely determined by the activation of cyclin-dependent kinases by regulatory protein subunits called cyclins, different forms of which are produced at each stage of the cell cycle to control the specific events that occur therein.

Rhodanese is a mitochondrial enzyme that detoxifies cyanide (CN−) by converting it to thiocyanate. In enzymatology, the common name is listed as thiosulfate sulfurtransferase. The diagram on the right shows the crystallographically-determined structure of rhodanese.

A cyclin-dependent kinase inhibitor protein(also known as CKIs, CDIs, or CDKIs) is a protein which inhibits the enzyme cyclin-dependent kinase (CDK) and Cyclin activity by stopping the cell cycle if there are unfavorable conditions, therefore, acting as tumor suppressors. Cell cycle progression is stopped by Cyclin-dependent kinase inhibitor protein at the G1 phase. CKIs are vital proteins within the control system that point out whether the process of DNA synthesis, mitosis, and cytokines control one another. If a malfunction prevents the successful completion of DNA synthesis during the G1 phase, a signal is sent to delay or stop the progression to the S phase. Cyclin-dependent kinase inhibitor proteins are essential in the regulation of the cell cycle. If cell mutations surpass the cell cycle checkpoints during cell cycle regulation, it can result in various types of cancer.

Cyclin-dependent kinase 1 also known as CDK1 or cell division cycle protein 2 homolog is a highly conserved protein that functions as a serine/threonine protein kinase, and is a key player in cell cycle regulation. It has been highly studied in the budding yeast S. cerevisiae, and the fission yeast S. pombe, where it is encoded by genes cdc28 and cdc2, respectively. With its cyclin partners, Cdk1 forms complexes that phosphorylate a variety of target substrates ; phosphorylation of these proteins leads to cell cycle progression.
G2/mitotic-specific cyclin-B1 is a protein that in humans is encoded by the CCNB1 gene.

M-phase inducer phosphatase 1 also known as dual specificity phosphatase Cdc25A is a protein that in humans is encoded by the cell division cycle 25 homolog A (CDC25A) gene.

M-phase inducer phosphatase 2 is an enzyme that in humans is encoded by the CDC25B gene.

M-phase inducer phosphatase 3 is an enzyme that in humans is encoded by the CDC25C gene.

Membrane-associated tyrosine- and threonine-specific cdc2-inhibitory kinase also known as Myt1 kinase is an enzyme that in humans is encoded by the PKMYT1 gene.

Wee1 is a nuclear kinase belonging to the Ser/Thr family of protein kinases in the fission yeast Schizosaccharomyces pombe. Wee1 has a molecular mass of 96 kDa and is a key regulator of cell cycle progression. It influences cell size by inhibiting the entry into mitosis, through inhibiting Cdk1. Wee1 has homologues in many other organisms, including mammals.
A series of biochemical switches control transitions between and within the various phases of the cell cycle. The cell cycle is a series of complex, ordered, sequential events that control how a single cell divides into two cells, and involves several different phases. The phases include the G1 and G2 phases, DNA replication or S phase, and the actual process of cell division, mitosis or M phase. During the M phase, the chromosomes separate and cytokinesis occurs.

The G2-M DNA damage checkpoint is an important cell cycle checkpoint in eukaryotic organisms that ensures that cells don't initiate mitosis until damaged or incompletely replicated DNA is sufficiently repaired. Cells with a defective G2-M checkpoint will undergo apoptosis or death after cell division if they enter the M phase before repairing their DNA. The defining biochemical feature of this checkpoint is the activation of M-phase cyclin-CDK complexes, which phosphorylate proteins that promote spindle assembly and bring the cell to metaphase.
References
- ↑ cdc25+ functions as an inducer in the mitotic control of fission yeast. Russell P, Nurse P. (1986 ) Cell: 45:145-53
- ↑ Boutros, R., Lobjois, V. & Ducommun, B. CDC25 phosphatases in cancer cells: key players? Good targets?. Nat Rev Cancer 7, 495–507 (2007). https://doi.org/10.1038/nrc2169
- ↑ Strausfeld U, Labbé JC, Fesquet D, et al. (May 1991). "Dephosphorylation and activation of a p34cdc2/cyclin B complex in vitro by human CDC25 protein". Nature. 351 (6323): 242–5. Bibcode:1991Natur.351..242S. doi:10.1038/351242a0. PMID 1828290. S2CID 4372756.
- ↑ Morgan, David. The Cell Cycle: Principles of Control. London: New Science Press, 2007. 96-98, 34-35. Print.
- 1 2 3 4 "Presentation on CDC25 PHOSPHATASES: A Potential Target for Novel Anticancer Agents". Archived from the original on 2016-03-03. Retrieved 2010-03-11.
- ↑ Sadhu K, Reed SI, Richardson H, Russell P (July 1990). "Human homolog of fission yeast cdc25 mitotic inducer is expressed predominantly in G2". Proc. Natl. Acad. Sci. U.S.A. 87 (13): 5139–43. doi: 10.1073/pnas.87.13.5139 . PMC 54277 . PMID 2195549.
- ↑ Landrieu I, da Costa M, De Veylder L, et al. (September 2004). "A small CDC25 dual-specificity tyrosine-phosphatase isoform in Arabidopsis thaliana". Proc. Natl. Acad. Sci. U.S.A. 101 (36): 13380–5. Bibcode:2004PNAS..10113380L. doi: 10.1073/pnas.0405248101 . PMC 516575 . PMID 15329414.
- ↑ Edgar BA, O'Farrell PH (April 1989). "Genetic Control of Cell Division Patterns in the Drosophila Embryo". Cell. 57 (1): 177–87. doi:10.1016/0092-8674(89)90183-9. PMC 2755076 . PMID 2702688.
- ↑ Alphey L, Jimenez J, White-Cooper H, Dawson I, Nurse P, Glover DM (June 1992). "twine, a cdc25 homolog that functions in the male and female germline of Drosophila". Cell. 69 (6): 977–88. doi:10.1016/0092-8674(92)90616-K. PMID 1606618. S2CID 29299686.
- ↑ Ashcroft NR, Kosinski ME, Wickramasinghe D, Donovan PJ, Golden A (July 1998). "The four cdc25 genes from the nematode Caenorhabditis elegans". Gene. 214 (1–2): 59–66. doi:10.1016/S0378-1119(98)00228-5. PMID 9651482.
- ↑ Ferguson AM, White LS, Donovan PJ, Piwnica-Worms H (April 2005). "Normal Cell Cycle and Checkpoint Responses in Mice and Cells Lacking Cdc25B and Cdc25C Protein Phosphatases". Mol. Cell. Biol. 25 (7): 2853–60. doi:10.1128/MCB.25.7.2853-2860.2005. PMC 1061644 . PMID 15767688.
- ↑ Kristjánsdóttir K, Rudolph J (August 2004). "Cdc25 phosphatases and cancer". Chem. Biol. 11 (8): 1043–51. doi: 10.1016/j.chembiol.2004.07.007 . PMID 15324805.