Walking is one of the main gaits of terrestrial locomotion among legged animals. Walking is typically slower than running and other gaits. Walking is defined by an "inverted pendulum" gait in which the body vaults over the stiff limb or limbs with each step. This applies regardless of the usable number of limbs—even arthropods, with six, eight, or more limbs, walk. In humans, walking has health benefits including improved mental health and reduced risk of cardiovascular disease and death.
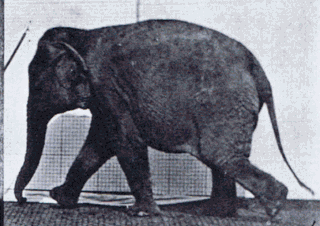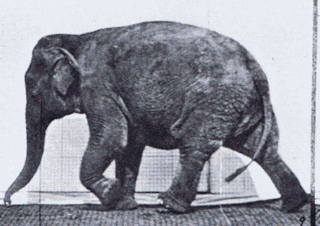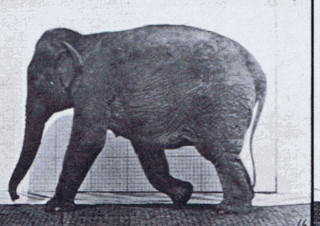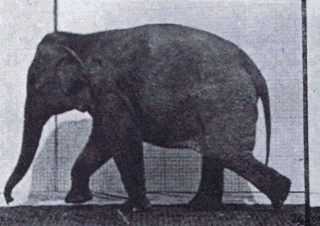
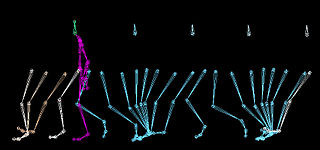
Gait is the pattern of movement of the limbs of animals, including humans, during locomotion over a solid substrate. Most animals use a variety of gaits, selecting gait based on speed, terrain, the need to maneuver, and energetic efficiency. Different animal species may use different gaits due to differences in anatomy that prevent use of certain gaits, or simply due to evolved innate preferences as a result of habitat differences. While various gaits are given specific names, the complexity of biological systems and interacting with the environment make these distinctions "fuzzy" at best. Gaits are typically classified according to footfall patterns, but recent studies often prefer definitions based on mechanics. The term typically does not refer to limb-based propulsion through fluid mediums such as water or air, but rather to propulsion across a solid substrate by generating reactive forces against it.

Gait analysis is the systematic study of animal locomotion, more specifically the study of human motion, using the eye and the brain of observers, augmented by instrumentation for measuring body movements, body mechanics, and the activity of the muscles. Gait analysis is used to assess and treat individuals with conditions affecting their ability to walk. It is also commonly used in sports biomechanics to help athletes run more efficiently and to identify posture-related or movement-related problems in people with injuries.

A gait is a manner of limb movements made during locomotion. Human gaits are the various ways in which humans can move, either naturally or as a result of specialized training. Human gait is defined as bipedal forward propulsion of the center of gravity of the human body, in which there are sinuous movements of different segments of the body with little energy spent. Varied gaits are characterized by differences such as limb movement patterns, overall velocity, forces, kinetic and potential energy cycles, and changes in contact with the ground.
Biarticular muscles are muscles that cross two joints rather than just one, such as the hamstrings which cross both the hip and the knee. The function of these muscles is complex and often depends upon both their anatomy and the activity of other muscles at the joints in question. Their role in movement is currently poorly understood.
In terrestrial animals, plantigrade locomotion means walking with the toes and metatarsals flat on the ground. It is one of three forms of locomotion adopted by terrestrial mammals. The other options are digitigrade, walking on the toes with the heel and wrist permanently raised, and unguligrade, walking on the nail or nails of the toes with the heel/wrist and the digits permanently raised. The leg of a plantigrade mammal includes the bones of the upper leg (femur/humerus) and lower leg. The leg of a digitigrade mammal also includes the metatarsals/metacarpals, the bones that in a human compose the arch of the foot and the palm of the hand. The leg of an unguligrade mammal also includes the phalanges, the finger and toe bones.

Animal locomotion, in ethology, is any of a variety of methods that animals use to move from one place to another. Some modes of locomotion are (initially) self-propelled, e.g., running, swimming, jumping, flying, hopping, soaring and gliding. There are also many animal species that depend on their environment for transportation, a type of mobility called passive locomotion, e.g., sailing, kiting (spiders), rolling or riding other animals (phoresis).
Robot locomotion is the collective name for the various methods that robots use to transport themselves from place to place.
In physiology, motor coordination is the orchestrated movement of multiple body parts as required to accomplish intended actions, like walking. This coordination is achieved by adjusting kinematic and kinetic parameters associated with each body part involved in the intended movement. The modifications of these parameters typically relies on sensory feedback from one or more sensory modalities, such as proprioception and vision.

Campaniform sensilla are a class of mechanoreceptors found in insects, which respond to local stress and strain within the animal's cuticle. Campaniform sensilla function as proprioceptors that detect mechanical load as resistance to muscle contraction, similar to mammalian Golgi tendon organs. Sensory feedback from campaniform sensilla is integrated in the control of posture and locomotion.
A facultative biped is an animal that is capable of walking or running on two legs (bipedal), as a response to exceptional circumstances (facultative), while normally walking or running on four limbs or more. In contrast, obligate bipedalism is where walking or running on two legs is the primary method of locomotion. Facultative bipedalism has been observed in several families of lizards and multiple species of primates, including sifakas, capuchin monkeys, baboons, gibbons, gorillas, bonobos and chimpanzees. Several dinosaur and other prehistoric archosaur species are facultative bipeds, most notably ornithopods and marginocephalians, with some recorded examples within sauropodomorpha. Different facultatively bipedal species employ different types of bipedalism corresponding to the varying reasons they have for engaging in facultative bipedalism. In primates, bipedalism is often associated with food gathering and transport. In lizards, it has been debated whether bipedal locomotion is an advantage for speed and energy conservation or whether it is governed solely by the mechanics of the acceleration and lizard's center of mass. Facultative bipedalism is often divided into high-speed (lizards) and low-speed (gibbons), but some species cannot be easily categorized into one of these two. Facultative bipedalism has also been observed in cockroaches and some desert rodents.

Arboreal locomotion is the locomotion of animals in trees. In habitats in which trees are present, animals have evolved to move in them. Some animals may scale trees only occasionally, but others are exclusively arboreal. The habitats pose numerous mechanical challenges to animals moving through them and lead to a variety of anatomical, behavioral and ecological consequences as well as variations throughout different species. Furthermore, many of these same principles may be applied to climbing without trees, such as on rock piles or mountains.

Comparative foot morphology involves comparing the form of distal limb structures of a variety of terrestrial vertebrates. Understanding the role that the foot plays for each type of organism must take account of the differences in body type, foot shape, arrangement of structures, loading conditions and other variables. However, similarities also exist among the feet of many different terrestrial vertebrates. The paw of the dog, the hoof of the horse, the manus (forefoot) and pes (hindfoot) of the elephant, and the foot of the human all share some common features of structure, organization and function. Their foot structures function as the load-transmission platform which is essential to balance, standing and types of locomotion.

Undulatory locomotion is the type of motion characterized by wave-like movement patterns that act to propel an animal forward. Examples of this type of gait include crawling in snakes, or swimming in the lamprey. Although this is typically the type of gait utilized by limbless animals, some creatures with limbs, such as the salamander, forgo use of their legs in certain environments and exhibit undulatory locomotion. In robotics this movement strategy is studied in order to create novel robotic devices capable of traversing a variety of environments.
Human locomotion is considered to take two primary forms: walking and running. In contrast, many quadrupeds have three distinct forms of locomotion: walk, trot, and gallop. Walking is a form of locomotion defined by a double support phase when both feet are on the ground at the same time. Running is a form of locomotion that does not have this double support phase.
X-ray motion analysis is a technique used to track the movement of objects using X-rays. This is done by placing the subject to be imaged in the center of the X-ray beam and recording the motion using an image intensifier and a high-speed camera, allowing for high quality videos sampled many times per second. Depending on the settings of the X-rays, this technique can visualize specific structures in an object, such as bones or cartilage. X-ray motion analysis can be used to perform gait analysis, analyze joint movement, or record the motion of bones obscured by soft tissue. The ability to measure skeletal motions is a key aspect to one's understanding of vertebrate biomechanics, energetics, and motor control.
A metachronal swimming or metachronal rowing is the swimming technique used by animals with multiple pairs of swimming legs. In this technique, appendages are sequentially stroked in a back-to-front wave moving along the animal’s body. In literature, while metachronal rhythm or metachronal wave usually refer to the movement of cilia; metachronal coordination, metachronal beating, metachronal swimming or metachronal rowing usually refer to the leg movement of arthropods, such as mantis shrimp, copepods, antarctic krill etc. though all of them refer to the similar locomotion pattern.
Hair-plates are a type of proprioceptor found in the folds of insect joints. They consist of a cluster of hairs, in which each hair is innervated by a single mechanosensory neuron. Functionally, hair-plates operate as "limit-detectors" by signaling the extreme ranges of motion of a joint.
Arachnid locomotion is the various means by which arachnids walk, run, or jump; they make use of more than muscle contraction, employing additional methods like hydraulic compression. Another adaptation seen especially in larger arachnid variants is inclusion of elastic connective tissues.

The femoral chordotonal organ is a group of mechanosensory neurons found in an insect leg that detects the movements and the position of the femur/tibia joint. It is thought to function as a proprioceptor that is critical for precise control of leg position by sending the information regarding the femur/tibia joint to the motor circuits in the ventral nerve cord and the brain


