Gavialidae is a family of large semiaquatic crocodilians with elongated, narrow snouts. Gavialidae consists of two living species, the gharial and the false gharial, both occurring in Asia. Many extinct members are known from a broader range, including the recently extinct Hanyusuchus. Gavialids are generally regarded as lacking the jaw strength to capture the large mammalian prey favoured by crocodiles and alligators of similar size so their thin snout is best used to catch fish, however the false gharial has been found to have a generalist diet with mature adults preying upon larger vertebrates, such as ungulates.

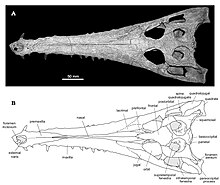
Toyotamaphimeia is a genus of extinct gavialid crocodylian which lived in Japan and Taiwan during the Pleistocene. A specimen recovered in 1964 at Osaka University during the construction of a new science building has been dated to around 430–380 thousand years old based on the stratum in which it was found. T. machikanensis was a fairly large crocodylian measuring approximately 6.3–7.3 metres (21–24 ft) long. Two species were named, T. machikanensis from Japan and T. taiwanicus from Taiwan, both originally described as members of the genus Tomistoma.

Gavialosuchus is an extinct genus of gavialoid crocodylian from the early Miocene of Europe. Currently only one species is recognized, as a few other species of Gavialosuchus have since been reclassified to other genera.
Argochampsa is an extinct genus of eusuchian crocodylomorph, usually regarded as a gavialoid crocodilian, related to modern gharials. It lived in the Paleocene of Morocco. Described by Hua and Jouve in 2004, the type species is A. krebsi, with the species named for Bernard Krebs. Argochampsa had a long narrow snout, and appears to have been marine in habits.

Harpacochampsa is a poorly known Early Miocene crocodilian from the Bullock Creek lagerstätte of the Northern Territory, Australia. The current specimen consists of a partial skull and fragments of a long, slender snout reminiscent of that of a false gharial, demonstrating that it was a piscivore in life.

Dollosuchus is an extinct monospecific genus of tomistomine crocodilian originally named as a species of Gavialis. It is a basal form possibly related to Kentisuchus, according to several phylogenetic analyses that have been conducted in recent years, and is the oldest known tomistomine to date. Fossils have been found from Belgium and the United Kingdom. It had large supratemporal fenestrae in relation to its orbits, similar to Kentisuchus and Thecachampsa.
Eogavialis is an extinct genus of eusuchian crocodylomorph, usually regarded as a gavialoid crocodylian. It superficially resembles Tomistoma schlegelii, the extant false gharial, and consequently material from the genus was originally referred to Tomistoma. Indeed, it was not until 1982 that the name Eogavialis was constructed after it was realised that the specimens were from a more basal form.
Eothoracosaurus is an extinct monospecific genus of eusuchian crocodylomorphs found in Eastern United States which existed during the Late Cretaceous period. Eothoracosaurus is considered to belong to an informally named clade called the "thoracosaurs", named after the closely related Thoracosaurus. Thoracosaurs in general were traditionally thought to be related to the modern false gharial, largely because the nasal bones contact the premaxillae, but phylogenetic work starting in the 1990s instead supported affinities within gavialoid exclusive of such forms. Even more recent phylogenetic studies suggest that thoracosaurs might instead be non-crocodilian eusuchians.

Gryposuchus is an extinct genus of gavialid crocodilian. Fossils have been found from Argentina, Colombia, Venezuela, Brazil and the Peruvian Amazon. The genus existed during the Miocene epoch. One recently described species, G. croizati, grew to an estimated length of 10 metres (33 ft). Gryposuchus is the type genus of the subfamily Gryposuchinae, although a 2018 study indicates that Gryposuchinae and Gryposuchus might be paraphyletic and rather an evolutionary grade towards the gharial.
Ikanogavialis is an extinct genus of gavialid crocodilian. Fossils have been found in the Urumaco Formation in Urumaco, Venezuela and the Solimões Formation of Brazil. The strata from which remains are found are late Miocene in age, rather than Pliocene as was once thought. A possible member of this genus survived into the Late Holocene on Muyua or Woodlark Island in Papua New Guinea.

Kentisuchus is an extinct genus of gavialoid crocodylian, traditionally regarded as a member of the subfamily Tomistominae. Fossils have been found from England and France that date back to the early Eocene. The genus has also been recorded from Ukraine, but it unclear whether specimens from Ukraine are referable to Kentisuchus.

Thoracosaurus is an extinct genus of long-snouted eusuchian which existed during the Late Cretaceous and Early Paleocene in North America and Europe.
Maroccosuchus zennaroi is an extinct gavialoid crocodylian from the Early Eocene of Morocco, traditionally regarded as a member of the subfamily Tomistominae.
Paratomistoma is an extinct monospecific genus of gavialoid crocodylian. It is based on the holotype specimen CGM 42188, a partial posterior skull and lower jaw discovered at Wadi Hitan, Egypt, in Middle Eocene-age rocks of the Gehannam Formation. The skull is unfused but considered morphologically mature. Paratomistoma was named in 2000 by Christopher Brochu and Philip Gingerich; the type species is P. courti in honor of Nicholas Court, who found CGM 42188. They performed a phylogenetic analysis and found Paratomistoma to be a derived member of Tomistominae, related to the false gharial. It may have been a marine or coastal crocodilian.
Prodiplocynodon is an extinct genus of basal crocodyloid crocodylian. It is one of the only crocodyloids known from the Cretaceous and existed during the Maastrichtian stage. The only species of Prodiplocynodon is the type species P. langi from the Lance Formation of Wyoming, known only from a single holotype skull lacking the lower jaw.

Thecachampsa is an extinct genus of gavialoid crocodylian, traditionally regarded as a member of the subfamily Tomistominae. Fossils have been found from the eastern United States in deposits of Miocene age. Those named in the 19th century were distinguished primarily by the shape of their teeth, and have since been combined with T. antiquus. More recently erected species were reassigned from other genera, although their assignment to Thecachampsa has since been questioned.
Gryposuchinae is an extinct subfamily of gavialid crocodylians. Gryposuchines lived mainly in the Miocene of South America. However, "Ikanogavialis" papuensis may have survived more recently, into the Late Pleistocene/Holocene. Most were long-snouted coastal forms. The group was named in 2007 and includes genera such as Gryposuchus and Aktiogavialis, although a 2018 study indicates that the group might be paraphyletic and rather an evolutionary grade towards the gharial.

Gavialoidea is one of three superfamilies of crocodylians, the other two being Alligatoroidea and Crocodyloidea. Although many extinct species are known, only the gharial Gavialis gangeticus and the false gharial Tomistoma schlegelii are alive today, with Hanyusuchus having become extinct in the last few centuries.

Gavialis browni is an extinct species of the crocodylian genus Gavialis and a close relative of the living gharial Gavialis gangeticus.
Tomistoma cairense is an extinct species of gavialoid crocodilian from the Lutetian stage of the Eocene era. It lived in North East Africa, especially Egypt. Remains of T. cairense have been found in the Mokattam Formation, in Mokattam, Egypt. Tomistoma cairense did not have a Maxilla process within their lacrimal gland, whereas all extant (living) crocodilians do.