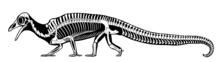
Archaeopteryx, sometimes referred to by its German name, "Urvogel", is a genus of avian dinosaurs. The name derives from the ancient Greek ἀρχαῖος (archaīos), meaning "ancient", and πτέρυξ (ptéryx), meaning "feather" or "wing". Between the late 19th century and the early 21st century, Archaeopteryx was generally accepted by palaeontologists and popular reference books as the oldest-known bird. Older potential avialans have since been identified, including Anchiornis, Xiaotingia, and Aurornis.

Theropoda, whose members are known as theropods, is a dinosaur clade that is characterized by hollow bones and three toes and claws on each limb. Theropods are generally classed as a group of saurischian dinosaurs. They were ancestrally carnivorous, although a number of theropod groups evolved to become herbivores and omnivores. Theropods first appeared during the Carnian age of the late Triassic period 231.4 million years ago (Ma) and included the majority of large terrestrial carnivores from the Early Jurassic until at least the close of the Cretaceous, about 66 Ma. In the Jurassic, birds evolved from small specialized coelurosaurian theropods, and are today represented by about 10,500 living species.

Sinosauropteryx is a compsognathid dinosaur. Described in 1996, it was the first dinosaur taxon outside of Avialae to be found with evidence of feathers. It was covered with a coat of very simple filament-like feathers. Structures that indicate colouration have also been preserved in some of its feathers, which makes Sinosauropteryx the first non-avialian dinosaurs where colouration has been determined. The colouration includes a reddish and light banded tail. Some contention has arisen with an alternative interpretation of the filamentous impression as remains of collagen fibres, but this has not been widely accepted.

Confuciusornis is a genus of basal crow-sized avialan from the Early Cretaceous Period of the Yixian and Jiufotang Formations of China, dating from 125 to 120 million years ago. Like modern birds, Confuciusornis had a toothless beak, but closer and later relatives of modern birds such as Hesperornis and Ichthyornis were toothed, indicating that the loss of teeth occurred convergently in Confuciusornis and living birds. It was thought to be the oldest known bird to have a beak, though this title now belongs to an earlier relative Eoconfuciusornis. It was named after the Chinese moral philosopher Confucius. Confuciusornis is one of the most abundant vertebrates found in the Yixian Formation, and several hundred complete specimens have been found.

Tetanurae is a clade that includes most theropod dinosaurs, including megalosauroids, allosauroids, tyrannosauroids, ornithomimosaurs, compsognathids and maniraptorans. Tetanurans are defined as all theropods more closely related to modern birds than to Ceratosaurus and contain the majority of predatory dinosaur diversity. Tetanurae likely diverged from its sister group, Ceratosauria, during the late Triassic. Tetanurae first appeared in the fossil record by the Early Jurassic about 190 mya and by the Middle Jurassic had become globally distributed.

Avimimus, meaning "bird mimic", is a genus of oviraptorosaurian theropod dinosaur, named for its bird-like characteristics, that lived in the late Cretaceous in what is now Mongolia, around 85 to 70 million years ago.

Postosuchus, meaning "Crocodile from Post", is an extinct genus of rauisuchid reptiles comprising two species, P. kirkpatricki and P. alisonae, that lived in what is now North America during the Late Triassic. Postosuchus is a member of the clade Pseudosuchia, the lineage of archosaurs that includes modern crocodilians. Its name refers to Post Quarry, a place in Texas where many fossils of the type species, P. kirkpatricki, were found. It was one of the apex predators of its area during the Triassic, larger than the small dinosaur predators of its time. It was a hunter which probably preyed on large bulky herbivores like dicynodonts and many other creatures smaller than itself.

Rahonavis is a genus of bird-like theropods from the Late Cretaceous of what is now northwestern Madagascar. It is known from a partial skeleton found by Catherine Forster and colleagues in Maevarano Formation rocks at a quarry near Berivotra, Mahajanga Province. Rahonavis was a small predator, at about 70 centimetres (2.3 ft) long and 0.45-2.27 kg, with the typical Velociraptor-like raised sickle claw on the second toe. It was originally the first African coelurosaur until the discovery of Nqwebasaurus in 2000.
Podokesaurus is a genus of coelophysoid dinosaur that lived in what is now the eastern United States during the Early Jurassic Period. The first fossil was discovered by the geologist Mignon Talbot near Mount Holyoke, Massachusetts, in 1910. The specimen was fragmentary, preserving much of the body, limbs, and tail. In 1911, Talbot described and named the new genus and species Podokesaurus holyokensis based on it. The full name can be translated as "swift-footed lizard of Holyoke". This discovery made Talbot the first woman to find and describe a non-bird dinosaur. The holotype fossil was recognized as significant and was studied by other researchers, but was lost when the building it was kept in burned down in 1917; no unequivocal Podokesaurus specimens have since been discovered. It was made state dinosaur of Massachusetts in 2022.

Marasuchus is a genus of basal dinosauriform archosaur which is possibly synonymous with Lagosuchus. Both genera lived during the Late Triassic in what is now La Rioja Province, Argentina. Marasuchus contains a single species, Marasuchus lilloensis.
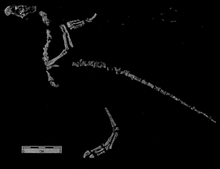
Shuvosaurus is a genus of beaked reptile from the Late Triassic of western Texas. Despite looking superficially similar to a theropod dinosaur, it is actually more closely related to crocodilians.The vertebrae of Gojirasaurus now are now known to belong to Shuvosaurus.

Sinosaurus is an extinct genus of theropod dinosaur which lived during the Early Jurassic Period. It was a bipedal carnivore approximately 5.5 metres (18 ft) in length and 300 kilograms (660 lb) in body mass. Fossils of the animal were found at the Lufeng Formation, in the Yunnan Province of China.
The evolution of birds began in the Jurassic Period, with the earliest birds derived from a clade of theropod dinosaurs named Paraves. Birds are categorized as a biological class, Aves. For more than a century, the small theropod dinosaur Archaeopteryx lithographica from the Late Jurassic period was considered to have been the earliest bird. Modern phylogenies place birds in the dinosaur clade Theropoda. According to the current consensus, Aves and a sister group, the order Crocodilia, together are the sole living members of an unranked reptile clade, the Archosauria. Four distinct lineages of bird survived the Cretaceous–Paleogene extinction event 66 million years ago, giving rise to ostriches and relatives (Palaeognathae), waterfowl (Anseriformes), ground-living fowl (Galliformes), and "modern birds" (Neoaves).

Poposaurus is an extinct genus of pseudosuchian archosaur from the Late Triassic of the southwestern United States. It belongs to the clade Poposauroidea, an unusual group of Triassic pseudosuchians that includes sail-backed, beaked, and aquatic forms. Fossils have been found in Wyoming, Utah, Arizona, and Texas. Except for the skull, most parts of the skeleton are known. The type species, P. gracilis, was described and named by Maurice Goldsmith Mehl in 1915. A second species, P. langstoni, was originally the type species of the genus Lythrosuchus. Since it was first described, Poposaurus has been variously classified as a dinosaur, a phytosaur, and a "rauisuchian".
The scientific question of within which larger group of animals birds evolved has traditionally been called the "origin of birds". The present scientific consensus is that birds are a group of maniraptoran theropod dinosaurs that originated during the Mesozoic Era.

John Alan Feduccia is a paleornithologist specializing in the origins and phylogeny of birds. He is S. K. Heninger Distinguished Professor Emeritus at the University of North Carolina. Feduccia's authored works include three major books, The Age of Birds, The Origin and Evolution of Birds, Riddle of the Feathered Dragons, and many peer-reviewed papers in ornithological and biological journals.
Zhongornis is a genus of primitive avialan that lived during the Early Cretaceous. It was found in rocks of the Yixian Formation in Lingyuan City (China), and described by Gao et al. in 2008.

Limusaurus is a genus of theropod dinosaur that lived in what is now China during the Late Jurassic, around 161 to 157 million years ago. The type and only species Limusaurus inextricabilis was described in 2009 from specimens found in the Upper Shishugou Formation in the Junggar Basin of China. The genus name consists of the Latin words for "mud" and "lizard", and the species name means "impossible to extricate", both referring to these specimens possibly dying after being mired. Limusaurus was a small, slender animal, about 1.7 m in length and 15 kg (33 lb) in weight, which had a long neck and legs but very small forelimbs. It underwent a drastic morphological transformation as it aged: while juveniles were toothed, these teeth were completely lost and replaced by a beak with age. Several of these features were convergently similar to the later ornithomimid theropods as well as the earlier non-dinosaurian shuvosaurids.
This glossary explains technical terms commonly employed in the description of dinosaur body fossils. Besides dinosaur-specific terms, it covers terms with wider usage, when these are of central importance in the study of dinosaurs or when their discussion in the context of dinosaurs is beneficial. The glossary does not cover ichnological and bone histological terms, nor does it cover measurements.

The Origin of Birds is an early synopsis of bird evolution written in 1926 by Gerhard Heilmann, a Danish artist and amateur zoologist. The book was born from a series of articles published between 1913 and 1916 in Danish, and although republished as a book it received mainly criticism from established scientists and got little attention within Denmark. The English edition of 1926, however, became highly influential at the time due to the breadth of evidence synthesized as well as the artwork used to support its arguments. It was considered the last word on the subject of bird evolution for several decades after its publication.