
An axon or nerve fiber is a long, slender projection of a nerve cell, or neuron, in vertebrates, that typically conducts electrical impulses known as action potentials away from the nerve cell body. The function of the axon is to transmit information to different neurons, muscles, and glands. In certain sensory neurons, such as those for touch and warmth, the axons are called afferent nerve fibers and the electrical impulse travels along these from the periphery to the cell body and from the cell body to the spinal cord along another branch of the same axon. Axon dysfunction can be the cause of many inherited and acquired neurological disorders that affect both the peripheral and central neurons. Nerve fibers are classed into three types – group A nerve fibers, group B nerve fibers, and group C nerve fibers. Groups A and B are myelinated, and group C are unmyelinated. These groups include both sensory fibers and motor fibers. Another classification groups only the sensory fibers as Type I, Type II, Type III, and Type IV.

Charcot–Marie–Tooth disease (CMT) is a hereditary motor and sensory neuropathy of the peripheral nervous system characterized by progressive loss of muscle tissue and touch sensation across various parts of the body. This disease is the most commonly inherited neurological disorder, affecting about one in 2,500 people. It is named after those who classically described it: the Frenchman Jean-Martin Charcot (1825–1893), his pupil Pierre Marie (1853–1940), and the Briton Howard Henry Tooth (1856–1925).

Nicotinamide adenine dinucleotide (NAD) is a coenzyme central to metabolism. Found in all living cells, NAD is called a dinucleotide because it consists of two nucleotides joined through their phosphate groups. One nucleotide contains an adenine nucleobase and the other, nicotinamide. NAD exists in two forms: an oxidized and reduced form, abbreviated as NAD+ and NADH (H for hydrogen), respectively.

Wallerian degeneration is an active process of degeneration that results when a nerve fiber is cut or crushed and the part of the axon distal to the injury degenerates. A related process of dying back or retrograde degeneration known as 'Wallerian-like degeneration' occurs in many neurodegenerative diseases, especially those where axonal transport is impaired such as ALS and Alzheimer's disease. Primary culture studies suggest that a failure to deliver sufficient quantities of the essential axonal protein NMNAT2 is a key initiating event.
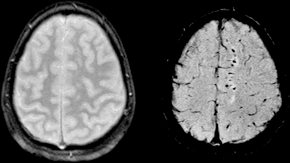
Diffuse axonal injury (DAI) is a brain injury in which scattered lesions occur over a widespread area in white matter tracts as well as grey matter. DAI is one of the most common and devastating types of traumatic brain injury and is a major cause of unconsciousness and persistent vegetative state after severe head trauma. It occurs in about half of all cases of severe head trauma and may be the primary damage that occurs in concussion. The outcome is frequently coma, with over 90% of patients with severe DAI never regaining consciousness. Those who awaken from the coma often remain significantly impaired.
Neurofilaments (NF) are classed as type IV intermediate filaments found in the cytoplasm of neurons. They are protein polymers measuring 10 nm in diameter and many micrometers in length. Together with microtubules (~25 nm) and microfilaments (7 nm), they form the neuronal cytoskeleton. They are believed to function primarily to provide structural support for axons and to regulate axon diameter, which influences nerve conduction velocity. The proteins that form neurofilaments are members of the intermediate filament protein family, which is divided into six types based on their gene organization and protein structure. Types I and II are the keratins which are expressed in epithelia. Type III contains the proteins vimentin, desmin, peripherin and glial fibrillary acidic protein (GFAP). Type IV consists of the neurofilament proteins NF-L, NF-M, NF-H and α-internexin. Type V consists of the nuclear lamins, and type VI consists of the protein nestin. The type IV intermediate filament genes all share two unique introns not found in other intermediate filament gene sequences, suggesting a common evolutionary origin from one primitive type IV gene.

CD38 (cluster of differentiation 38), also known as cyclic ADP ribose hydrolase is a glycoprotein found on the surface of many immune cells (white blood cells), including CD4+, CD8+, B lymphocytes and natural killer cells. CD38 also functions in cell adhesion, signal transduction and calcium signaling.

Cyclic ADP-ribose, frequently abbreviated as cADPR, is a cyclic adenine nucleotide (like cAMP) with two phosphate groups present on 5' OH of the adenosine (like ADP), further connected to another ribose at the 5' position, which, in turn, closes the cycle by glycosidic bonding to the nitrogen 1 (N1) of the same adenine base (whose position N9 has the glycosidic bond to the other ribose). The N1-glycosidic bond to adenine is what distinguishes cADPR from ADP-ribose (ADPR), the non-cyclic analog. cADPR is produced from nicotinamide adenine dinucleotide (NAD+) by ADP-ribosyl cyclases (EC 3.2.2.5) as part of a second messenger system.
Neuroregeneration involves the regrowth or repair of nervous tissues, cells or cell products. Neuroregenerative mechanisms may include generation of new neurons, glia, axons, myelin, or synapses. Neuroregeneration differs between the peripheral nervous system (PNS) and the central nervous system (CNS) by the functional mechanisms involved, especially in the extent and speed of repair. When an axon is damaged, the distal segment undergoes Wallerian degeneration, losing its myelin sheath. The proximal segment can either die by apoptosis or undergo the chromatolytic reaction, which is an attempt at repair. In the CNS, synaptic stripping occurs as glial foot processes invade the dead synapse.

Mitofusin-2 is a protein that in humans is encoded by the MFN2 gene. Mitofusins are GTPases embedded in the outer membrane of the mitochondria. In mammals MFN1 and MFN2 are essential for mitochondrial fusion. In addition to the mitofusins, OPA1 regulates inner mitochondrial membrane fusion, and DRP1 is responsible for mitochondrial fission.

Nicotinamide phosphoribosyltransferase, formerly known as pre-B-cell colony-enhancing factor 1 (PBEF1) or visfatin for its extracellular form (eNAMPT), is an enzyme that in humans is encoded by the NAMPT gene. The intracellular form of this protein (iNAMPT) is the rate-limiting enzyme in the nicotinamide adenine dinucleotide (NAD+) salvage pathway that converts nicotinamide to nicotinamide mononucleotide (NMN) which is responsible for most of the NAD+ formation in mammals. iNAMPT can also catalyze the synthesis of NMN from phosphoribosyl pyrophosphate (PRPP) when ATP is present. eNAMPT has been reported to be a cytokine (PBEF) that activates TLR4, that promotes B cell maturation, and that inhibits neutrophil apoptosis.

In enzymology, nicotinamide-nucleotide adenylyltransferase (NMNAT) (EC 2.7.7.1) are enzymes that catalyzes the chemical reaction

Nicotinamide mononucleotide adenylyltransferase 1 (NMNAT1) is an enzyme that in humans is encoded by the nmnat1 gene. It is a member of the nicotinamide-nucleotide adenylyltransferases (NMNATs) which catalyze nicotinamide adenine dinucleotide (NAD) synthesis.
Nicotinamide mononucleotide adenylyltransferase 2 (NMNAT2) is an enzyme that in humans is encoded by the NMNAT2 gene.

Ninjurin-1 is a protein that in humans is encoded by the NINJ1 gene.
Sirtuin-activating compounds (STAC) are chemical compounds having an effect on sirtuins, a group of enzymes that use NAD+ to remove acetyl groups from proteins. They are caloric restriction mimetic compounds that may be helpful in treating various aging-related diseases.

The toll-interleukin-1 receptor (TIR) homology domain is an intracellular signaling domain found in MyD88, SARM1, interleukin-1 receptors, toll receptors and many plant R proteins. It contains three highly conserved regions, and mediates protein-protein interactions between the toll-like receptors (TLRs) and signal-transduction components. TIR-like motifs are also found in plant proteins where they are involved in resistance to disease and in bacteria where they are associated with virulence. When activated, TIR domains recruit cytoplasmic adaptor proteins MyD88 (UniProt Q99836) and TOLLIP (toll-interacting protein, UniProt Q9H0E2). In turn, these associate with various kinases to set off signaling cascades. Some TIR domains have also been found to have intrinsic NAD+ cleavage activity, such as in SARM1. In the case of SARM1, the TIR NADase activity leads to the production of Nam, ADPR and cADPR and the activation of downstream pathways involved in Wallerian degeneration and neuron death.
Members of the very wide interleukin-1 receptor (IL-1R) family are characterized by extracellular immunoglobulin-like domains and intracellular Toll/Interleukin-1R (TIR) domain. It is a group of structurally homologous proteins, conserved throughout the species as it was identified from plants to mammals. Proteins of this family play important role in host defence, injury and stress. There are four main groups of TIR domain-containing proteins in animals; Toll-like receptors, Interleukin-1 receptor (IL-1R), cytosolic adaptor proteins and insect and nematode Toll. Each of these groups is involved mainly in host defence; Toll receptors are also involved in embryogenesis.
Preferential motor reinnervation (PMR) refers to the tendency of a regenerating axon in the peripheral nervous system (PNS) to reinnervate a motor pathway as opposed to a somatosensory pathway. PMR affects how nerves regenerate and reinnervate within the PNS after surgical procedures or traumatic injuries. It is important to understand in order to further develop axonal regrowth surgical techniques. Further research of preferential motor reinnervation will lead to a better understanding of peripheral nervous system function in the human body regarding cell roles and abilities.

Nicotinamide mononucleotide is a nucleotide derived from ribose, nicotinamide, nicotinamide riboside and niacin. In humans, several enzymes use NMN to generate nicotinamide adenine dinucleotide (NADH). In mice, it has been proposed that NMN is absorbed via the small intestine within 10 minutes of oral uptake and converted to nicotinamide adenine dinucleotide (NAD+) through the Slc12a8 transporter. However, this observation has been challenged, and the matter remains unsettled.



