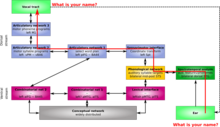
The visual cortex of the brain is the area of the cerebral cortex that processes visual information. It is located in the occipital lobe. Sensory input originating from the eyes travels through the lateral geniculate nucleus in the thalamus and then reaches the visual cortex. The area of the visual cortex that receives the sensory input from the lateral geniculate nucleus is the primary visual cortex, also known as visual area 1 (V1), Brodmann area 17, or the striate cortex. The extrastriate areas consist of visual areas 2, 3, 4, and 5.

The parietal lobe is one of the four major lobes of the cerebral cortex in the brain of mammals. The parietal lobe is positioned above the temporal lobe and behind the frontal lobe and central sulcus.
The temporal lobe is one of the four major lobes of the cerebral cortex in the brain of mammals. The temporal lobe is located beneath the lateral fissure on both cerebral hemispheres of the mammalian brain.

Wernicke's area, also called Wernicke's speech area, is one of the two parts of the cerebral cortex that are linked to speech, the other being Broca's area. It is involved in the comprehension of written and spoken language, in contrast to Broca's area, which is primarily involved in the production of language. It is traditionally thought to reside in Brodmann area 22, which is located in the superior temporal gyrus in the dominant cerebral hemisphere, which is the left hemisphere in about 95% of right-handed individuals and 70% of left-handed individuals.

Visual memory describes the relationship between perceptual processing and the encoding, storage and retrieval of the resulting neural representations. Visual memory occurs over a broad time range spanning from eye movements to years in order to visually navigate to a previously visited location. Visual memory is a form of memory which preserves some characteristics of our senses pertaining to visual experience. We are able to place in memory visual information which resembles objects, places, animals or people in a mental image. The experience of visual memory is also referred to as the mind's eye through which we can retrieve from our memory a mental image of original objects, places, animals or people. Visual memory is one of several cognitive systems, which are all interconnected parts that combine to form the human memory. Types of palinopsia, the persistence or recurrence of a visual image after the stimulus has been removed, is a dysfunction of visual memory.
Simultanagnosia is a rare neurological disorder characterized by the inability of an individual to perceive more than a single object at a time. This type of visual attention problem is one of three major components of Bálint's syndrome, an uncommon and incompletely understood variety of severe neuropsychological impairments involving space representation. The term "simultanagnosia" was first coined in 1924 by Wolpert to describe a condition where the affected individual could see individual details of a complex scene but failed to grasp the overall meaning of the image.

In psycholinguistics, language processing refers to the way humans use words to communicate ideas and feelings, and how such communications are processed and understood. Language processing is considered to be a uniquely human ability that is not produced with the same grammatical understanding or systematicity in even human's closest primate relatives.
Visual agnosia is an impairment in recognition of visually presented objects. It is not due to a deficit in vision, language, memory, or intellect. While cortical blindness results from lesions to primary visual cortex, visual agnosia is often due to damage to more anterior cortex such as the posterior occipital and/or temporal lobe(s) in the brain.[2] There are two types of visual agnosia: apperceptive agnosia and associative agnosia.

The inferior temporal gyrus is one of three gyri of the temporal lobe and is located below the middle temporal gyrus, connected behind with the inferior occipital gyrus; it also extends around the infero-lateral border on to the inferior surface of the temporal lobe, where it is limited by the inferior sulcus. This region is one of the higher levels of the ventral stream of visual processing, associated with the representation of objects, places, faces, and colors. It may also be involved in face perception, and in the recognition of numbers and words.
The perirhinal cortex is a cortical region in the medial temporal lobe that is made up of Brodmann areas 35 and 36. It receives highly processed sensory information from all sensory regions, and is generally accepted to be an important region for memory. It is bordered caudally by postrhinal cortex or parahippocampal cortex and ventrally and medially by entorhinal cortex.
Vision for perception and vision for action in neuroscience literature refers to two types of visual processing in the brain: visual processing to obtain information about the features of objects such as color, size, shape versus processing needed to guide movements such as catching a baseball. An idea is currently debated that these types of processing are done by anatomically different brain networks. Ventral visual stream subserves vision for perception, whereas dorsal visual stream subserves vision for action. This idea finds support in clinical research and animal experiments.
Auditory agnosia is a form of agnosia that manifests itself primarily in the inability to recognize or differentiate between sounds. It is not a defect of the ear or "hearing", but rather a neurological inability of the brain to process sound meaning. While auditory agnosia impairs the understanding of sounds, other abilities such as reading, writing, and speaking are not hindered. It is caused by bilateral damage to the anterior superior temporal gyrus, which is part of the auditory pathway responsible for sound recognition, the auditory "what" pathway.
The neuroanatomy of memory encompasses a wide variety of anatomical structures in the brain.
Recognition memory, a subcategory of explicit memory, is the ability to recognize previously encountered events, objects, or people. When the previously experienced event is reexperienced, this environmental content is matched to stored memory representations, eliciting matching signals. As first established by psychology experiments in the 1970s, recognition memory for pictures is quite remarkable: humans can remember thousands of images at high accuracy after seeing each only once and only for a few seconds.
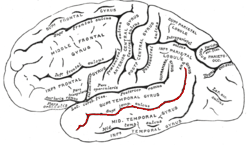
In the human brain, the superior temporal sulcus (STS) is the sulcus separating the superior temporal gyrus from the middle temporal gyrus in the temporal lobe of the brain. A sulcus is a deep groove that curves into the largest part of the brain, the cerebrum, and a gyrus is a ridge that curves outward of the cerebrum.
Visual object recognition refers to the ability to identify the objects in view based on visual input. One important signature of visual object recognition is "object invariance", or the ability to identify objects across changes in the detailed context in which objects are viewed, including changes in illumination, object pose, and background context.

A parasol cell, sometimes called an M cell or M ganglion cell, is one type of retinal ganglion cell (RGC) located in the ganglion cell layer of the retina. These cells project to magnocellular cells in the lateral geniculate nucleus (LGN) as part of the magnocellular pathway in the visual system. They have large cell bodies as well as extensive branching dendrite networks and as such have large receptive fields. Relative to other RGCs, they have fast conduction velocities. While they do show clear center-surround antagonism, they receive no information about color. Parasol ganglion cells contribute information about the motion and depth of objects to the visual system.
Phonagnosia is a type of agnosia, or loss of knowledge, that involves a disturbance in the recognition of familiar voices and the impairment of voice discrimination abilities in which the affected individual does not suffer from comprehension deficits. Phonagnosia is an auditory agnosia, an acquired auditory processing disorder resulting from brain damage, other auditory agnosias include cortical deafness and auditory verbal agnosia also known as pure word deafness.
Constructional apraxia is a neurological disorder in which people are unable to perform tasks or movements even though they understand the task, are willing to complete it, and have the physical ability to perform the movements. It is characterized by an inability or difficulty to build, assemble, or draw objects. Constructional apraxia may be caused by lesions in the parietal lobe following stroke or it may serve as an indicator for Alzheimer's disease.

Melvyn Alan Goodale FRSC, FRS is a Canadian neuroscientist. He was the founding Director of the Brain and Mind Institute at the University of Western Ontario where he holds the Canada Research Chair in Visual Neuroscience. He holds appointments in the Departments of Psychology, Physiology & Pharmacology, and Ophthalmology at Western. Goodale's research focuses on the neural substrates of visual perception and visuomotor control.