Y-chromosomal Aaron is the name given to the hypothesized most recent common ancestor of the patrilineal Jewish priestly caste known as Kohanim. According to the traditional understanding of the Hebrew Bible, this ancestor was Aaron, the brother of Moses.

Haplogroup G (M201) is a human Y-chromosome haplogroup. It is one of two branches of the parent haplogroup GHIJK, the other being HIJK.

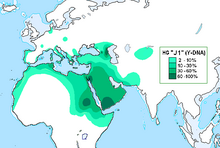
In human genetics, Haplogroup J-M172 or J2 is a Y-chromosome haplogroup which is a subclade (branch) of haplogroup J-M304. Haplogroup J-M172 is common in modern populations in Western Asia, Central Asia, South Asia, Southern Europe, Northwestern Iran and North Africa. It is thought that J-M172 may have originated between the Caucasus, Anatolia and/or Western Iran.

Haplogroup J-M304, also known as J, is a human Y-chromosome DNA haplogroup. It is believed to have evolved in Western Asia. The clade spread from there during the Neolithic, primarily into North Africa, the Horn of Africa, the Socotra Archipelago, the Caucasus, Europe, Anatolia, Central Asia, South Asia, and Southeast Asia.

E-M215 or E1b1b, formely known as E3b, is a major human Y-chromosome DNA haplogroup. E-M215 has two basal branches, E-M35 and E-M281. E-M35 is primarily distributed in North Africa and the Horn of Africa, and occurs at moderate frequencies in the Middle East, Europe, and Southern Africa. E-M281 occurs at a low frequency in Ethiopia.
Haplogroup E-M96 is a human Y-chromosome DNA haplogroup. It is one of the two main branches of the older and ancestral haplogroup DE, the other main branch being haplogroup D. The E-M96 clade is divided into two main subclades: the more common E-P147, and the less common E-M75.
Haplogroup K or K-M9 is a genetic lineage within human Y-chromosome DNA haplogroup. A sublineage of haplogroup IJK, K-M9, and its descendant clades represent a geographically widespread and diverse haplogroup. The lineages have long been found among males on every continent except Antarctica.

In human genetics, a human Y-chromosome DNA haplogroup is a haplogroup defined by mutations in the non-recombining portions of DNA from the male-specific Y chromosome. Many people within a haplogroup share similar numbers of short tandem repeats (STRs) and types of mutations called single-nucleotide polymorphisms (SNPs).
Haplogroup R, or R-M207, is a Y-chromosome DNA haplogroup. It is both numerous and widespread amongst modern populations.

Haplogroup T-M184, also known as Haplogroup T, is a human Y-chromosome DNA haplogroup. The unique-event polymorphism that defines this clade is the single-nucleotide polymorphism known as M184.

Haplogroup S-M230, also known as S1a1b, is a Y-chromosome DNA haplogroup. It is by far the most numerically significant subclade of Haplogroup S1a.
Haplogroup E-P147 is a human Y-chromosome DNA haplogroup. Haplogroup E-P147, along with the less common haplogroup E-M75, is one of the two main branches of the older haplogroup E-M96. The E-P147 clade is commonly observed throughout Africa and is divided into two subclades: the less common, haplogroup E-M132, and the more common, haplogroup E-P177.
Haplogroup E-M132, formerly known as E-M33 (E1a), is a human Y-chromosome DNA haplogroup. Along with E-P177, it is one of the two main branches of the older E-P147 paternal clade. E-M132 is divided into two primary sub-branches, E-M44 and E-Z958, with many descendant subclades.
Haplogroup E-P177 is a human Y-chromosome DNA haplogroup. E-P177 has two known subclades, which are haplogroup E-P2 and haplogroup E-P75.
Haplogroup E-V68, also known as E1b1b1a, is a major human Y-chromosome DNA haplogroup found in North Africa, the Horn of Africa, Western Asia and Europe. It is a subclade of the larger and older haplogroup, known as E1b1b or E-M215. The E1b1b1a lineage is identified by the presence of a single nucleotide polymorphism (SNP) mutation on the Y chromosome, which is known as V68. It is a subject of discussion and study in genetics as well as genetic genealogy, archaeology, and historical linguistics.
In human population genetics, Y-Chromosome haplogroups define the major lineages of direct paternal (male) lines back to a shared common ancestor in Africa. Men in the same haplogroup share a set of differences, or markers, on their Y-Chromosome, which distinguish them from men in other haplogroups. These UEPs, or markers used to define haplogroups, are SNP mutations. Y-Chromosome Haplogroups all form "family trees" or "phylogenies", with both branches or sub-clades diverging from a common haplogroup ancestor, and also with all haplogroups themselves linked into one family tree which traces back ultimately to the most recent shared male line ancestor of all men alive today, called in popular science Y Chromosome Adam.
Y-DNA haplogroups in populations of Europe are haplogroups of the male Y-chromosome found in European populations.
E-Z827, also known as E1b1b1b, is a major human Y-chromosome DNA haplogroup. It is the parent lineage to the E-Z830 and E-V257 subclades, and defines their common phylogeny. The former is predominantly found in the Middle East; the latter is most frequently observed in North Africa, with its E-M81 subclade observed among the ancient Guanche natives of the Canary Islands. E-Z827 is also found at lower frequencies in Europe, and in isolated parts of Southeast Africa.

Haplogroup R-M269 is the sub-clade of human Y-chromosome haplogroup R1b that is defined by the SNP marker M269. According to ISOGG 2020 it is phylogenetically classified as R1b1a1b. It underwent intensive research and was previously classified as R1b1a2, R1b1c, R1b1b2 and R1b1a1a2.

Haplogroup E-M2, also known as E1b1a1-M2, is a human Y-chromosome DNA haplogroup. E-M2 is primarily distributed within Africa followed by West Asia. More specifically, E-M2 is the predominant subclade in West Africa, Central Africa, Southern Africa, and the region of the African Great Lakes; it also occurs at moderate frequencies in North Africa, and the Middle East. E-M2 has several subclades, but many of these subhaplogroups are included in either E-L485 or E-U175. E-M2 is especially common among indigenous Africans who speak Niger-Congo languages, and was spread to Southern Africa and East Africa through the Bantu expansion.