
The thyroid, or thyroid gland, is an endocrine gland in vertebrates. In humans, it is in the neck and consists of two connected lobes. The lower two thirds of the lobes are connected by a thin band of tissue called the isthmus (pl.: isthmi). The thyroid gland is a butterfly-shaped gland located in the neck below the Adam's apple. Microscopically, the functional unit of the thyroid gland is the spherical thyroid follicle, lined with follicular cells (thyrocytes), and occasional parafollicular cells that surround a lumen containing colloid. The thyroid gland secretes three hormones: the two thyroid hormones – triiodothyronine (T3) and thyroxine (T4) – and a peptide hormone, calcitonin. The thyroid hormones influence the metabolic rate and protein synthesis and growth and development in children. Calcitonin plays a role in calcium homeostasis. Secretion of the two thyroid hormones is regulated by thyroid-stimulating hormone (TSH), which is secreted from the anterior pituitary gland. TSH is regulated by thyrotropin-releasing hormone (TRH), which is produced by the hypothalamus.
Hypothyroidism is a disorder of the endocrine system in which the thyroid gland does not produce enough thyroid hormones. It can cause a number of symptoms, such as poor ability to tolerate cold, a feeling of tiredness, constipation, slow heart rate, depression, and weight gain. Occasionally there may be swelling of the front part of the neck due to goitre. Untreated cases of hypothyroidism during pregnancy can lead to delays in growth and intellectual development in the baby or congenital iodine deficiency syndrome.
Thyroid-stimulating hormone (also known as thyrotropin, thyrotropic hormone, or abbreviated TSH) is a pituitary hormone that stimulates the thyroid gland to produce thyroxine (T4), and then triiodothyronine (T3) which stimulates the metabolism of almost every tissue in the body. It is a glycoprotein hormone produced by thyrotrope cells in the anterior pituitary gland, which regulates the endocrine function of the thyroid.
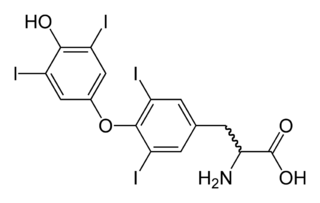
Triiodothyronine, also known as T3, is a thyroid hormone. It affects almost every physiological process in the body, including growth and development, metabolism, body temperature, and heart rate.

Levothyroxine, also known as L-thyroxine, is a synthetic form of the thyroid hormone thyroxine (T4). It is used to treat thyroid hormone deficiency (hypothyroidism), including a severe form known as myxedema coma. It may also be used to treat and prevent certain types of thyroid tumors. It is not indicated for weight loss. Levothyroxine is taken orally (by mouth) or given by intravenous injection. Levothyroxine has a half-life of 7.5 days when taken daily, so about six weeks is required for it to reach a steady level in the blood.

Propylthiouracil (PTU) is a medication used to treat hyperthyroidism. This includes hyperthyroidism due to Graves' disease and toxic multinodular goiter. In a thyrotoxic crisis it is generally more effective than methimazole. Otherwise it is typically only used when methimazole, surgery, and radioactive iodine is not possible. It is taken by mouth.
Thyroid function tests (TFTs) is a collective term for blood tests used to check the function of the thyroid. TFTs may be requested if a patient is thought to suffer from hyperthyroidism or hypothyroidism, or to monitor the effectiveness of either thyroid-suppression or hormone replacement therapy. It is also requested routinely in conditions linked to thyroid disease, such as atrial fibrillation and anxiety disorder.

Thyroxine 5-deiodinase also known as type III iodothyronine deiodinase (EC number 1.21.99.3) is an enzyme that in humans is encoded by the DIO3 gene. This enzyme catalyses the following chemical reaction
Desiccated thyroid, also known as thyroid extract, is thyroid gland that has been dried and powdered for medical use. It is used to treat hypothyroidism. It is less preferred than levothyroxine. It is taken by mouth. Maximal effects may take up to three weeks to occur.

The hypothalamic–pituitary–thyroid axis is part of the neuroendocrine system responsible for the regulation of metabolism and also responds to stress.

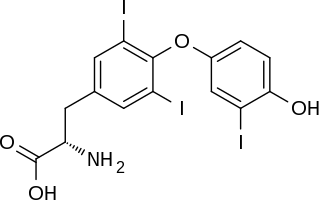
Reverse triiodothyronine (3,3′,5′-triiodothyronine, reverse T3, or rT3) is an isomer of triiodothyronine (3,5,3′ triiodothyronine, T3).
An antithyroid agent is a hormone inhibitor acting upon thyroid hormones.
Euthyroid sick syndrome (ESS) is a state of adaptation or dysregulation of thyrotropic feedback control wherein the levels of T3 and/or T4 are abnormal, but the thyroid gland does not appear to be dysfunctional. This condition may result from allostatic responses of hypothalamus-pituitary-thyroid feedback control, dyshomeostatic disorders, drug interferences, and impaired assay characteristics in critical illness.
Type II iodothyronine deiodinase is an enzyme that in humans is encoded by the DIO2 gene.
Thyroid hormones are any hormones produced and released by the thyroid gland, namely triiodothyronine (T3) and thyroxine (T4). They are tyrosine-based hormones that are primarily responsible for regulation of metabolism. T3 and T4 are partially composed of iodine, derived from food. A deficiency of iodine leads to decreased production of T3 and T4, enlarges the thyroid tissue and will cause the disease known as simple goitre.

Iopanoic acid is an iodine-containing radiocontrast medium used in cholecystography. Both iopanoic acid and ipodate sodium are potent inhibitors of thyroid hormone release from thyroid gland, as well as of peripheral conversion of thyroxine (T4) to triiodothyronine (T3). These compounds inhibit 5'deiodinase (5'DID-1 and 5'DID-2) enzymes, which catalyse T4-T3 conversion in the thyroid cell, liver, kidney, skeletal muscle, heart, brain, pituitary. This accounts for the dramatic improvement in both subjective and objective symptoms of hyperthyroidism, particularly when they are used as an adjunctive therapy with thioamides (propylthiouracil, carbimazole). They can be used in the treatment of patients with severe thyrotoxicosis (thyroid storm) and significant morbidity (e.g., myocardial infarction, or stroke) for rapid control of elevated plasma triiodothyronine concentrations. The use of iopanoic acid for treatment of thyrotoxicosis has been discontinued in the United States.

Iodotyrosine deiodinase, also known as iodotyrosine dehalogenase 1, is a type of deiodinase enzyme that scavenges iodide by removing it from iodinated tyrosine residues in the thyroid gland. These iodinated tyrosines are produced during thyroid hormone biosynthesis. The iodide that is scavenged by iodotyrosine deiodinase is necessary to again synthesize the thyroid hormones. After synthesis, the thyroid hormones circulate through the body to regulate metabolic rate, protein expression, and body temperature. Iodotyrosine deiodinase is thus necessary to keep levels of both iodide and thyroid hormones in balance.
Deiodinase (monodeiodinase) is a peroxidase enzyme that is involved in the activation or deactivation of thyroid hormones.
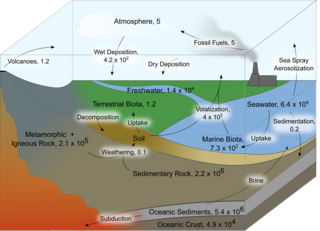
Iodine is an essential trace element in biological systems. It has the distinction of being the heaviest element commonly needed by living organisms as well as the second-heaviest known to be used by any form of life. It is a component of biochemical pathways in organisms from all biological kingdoms, suggesting its fundamental significance throughout the evolutionary history of life.
The sum activity of peripheral deiodinases is the maximum amount of triiodothyronine produced per time-unit under conditions of substrate saturation. It is assumed to reflect the activity of deiodinases outside the central nervous system and other isolated compartments. GD is therefore expected to reflect predominantly the activity of type I deiodinase.


