Related Research Articles

The mesoderm is the middle layer of the three germ layers that develops during gastrulation in the very early development of the embryo of most animals. The outer layer is the ectoderm, and the inner layer is the endoderm.
Segmentation in biology is the division of some animal and plant body plans into a linear series of repetitive segments that may or may not be interconnected to each other. This article focuses on the segmentation of animal body plans, specifically using the examples of the taxa Arthropoda, Chordata, and Annelida. These three groups form segments by using a "growth zone" to direct and define the segments. While all three have a generally segmented body plan and use a growth zone, they use different mechanisms for generating this patterning. Even within these groups, different organisms have different mechanisms for segmenting the body. Segmentation of the body plan is important for allowing free movement and development of certain body parts. It also allows for regeneration in specific individuals.

The somites are a set of bilaterally paired blocks of paraxial mesoderm that form in the embryonic stage of somitogenesis, along the head-to-tail axis in segmented animals. In vertebrates, somites subdivide into the dermatomes, myotomes, sclerotomes and syndetomes that give rise to the vertebrae of the vertebral column, rib cage, part of the occipital bone, skeletal muscle, cartilage, tendons, and skin.
Somitogenesis is the process by which somites form. Somites are bilaterally paired blocks of paraxial mesoderm that form along the anterior-posterior axis of the developing embryo in segmented animals. In vertebrates, somites give rise to skeletal muscle, cartilage, tendons, endothelium, and dermis.
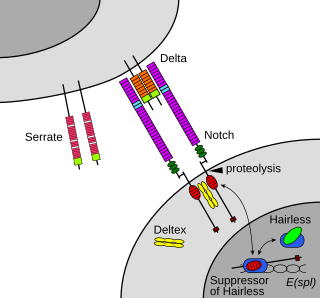
The Notch signaling pathway is a highly conserved cell signaling system present in most animals. Mammals possess four different notch receptors, referred to as NOTCH1, NOTCH2, NOTCH3, and NOTCH4. The notch receptor is a single-pass transmembrane receptor protein. It is a hetero-oligomer composed of a large extracellular portion, which associates in a calcium-dependent, non-covalent interaction with a smaller piece of the notch protein composed of a short extracellular region, a single transmembrane-pass, and a small intracellular region.
Organogenesis is the phase of embryonic development that starts at the end of gastrulation and continues until birth. During organogenesis, the three germ layers formed from gastrulation form the internal organs of the organism.

Intermediate mesoderm or intermediate mesenchyme is a narrow section of the mesoderm located between the paraxial mesoderm and the lateral plate of the developing embryo. The intermediate mesoderm develops into vital parts of the urogenital system.

Paraxial mesoderm, also known as presomitic or somitic mesoderm, is the area of mesoderm in the neurulating embryo that flanks and forms simultaneously with the neural tube. The cells of this region give rise to somites, blocks of tissue running along both sides of the neural tube, which form muscle and the tissues of the back, including connective tissue and the dermis.
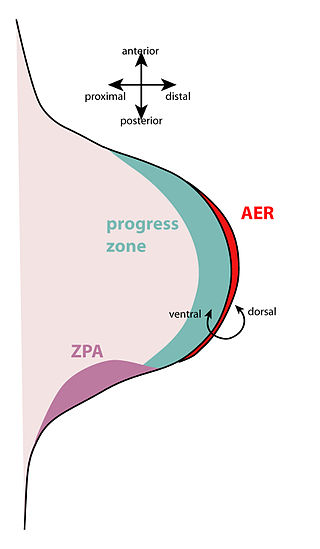
The apical ectodermal ridge (AER) is a structure that forms from the ectodermal cells at the distal end of each limb bud and acts as a major signaling center to ensure proper development of a limb. After the limb bud induces AER formation, the AER and limb mesenchyme—including the zone of polarizing activity (ZPA)—continue to communicate with each other to direct further limb development.
The limb bud is a structure formed early in vertebrate limb development. As a result of interactions between the ectoderm and underlying mesoderm, formation occurs roughly around the fourth week of development. In the development of the human embryo the upper limb bud appears in the third week and the lower limb bud appears four days later.

Otic vesicle, or auditory vesicle, consists of either of the two sac-like invaginations formed and subsequently closed off during embryonic development. It is part of the neural ectoderm, which will develop into the membranous labyrinth of the inner ear. This labyrinth is a continuous epithelium, giving rise to the vestibular system and auditory components of the inner ear. During the earlier stages of embryogenesis, the otic placode invaginates to produce the otic cup. Thereafter, the otic cup closes off, creating the otic vesicle. Once formed, the otic vesicle will reside next to the neural tube medially, and on the lateral side will be paraxial mesoderm. Neural crest cells will migrate rostral and caudal to the placode.
The scleraxis protein is a member of the basic helix-loop-helix (bHLH) superfamily of transcription factors. Currently two genes have been identified to code for identical scleraxis proteins.

Protein Wnt-3a is a protein that in humans is encoded by the WNT3A gene.

Beta-1,3-N-acetylglucosaminyltransferase lunatic fringe, (Lunatic Fringe), is a protein encoded in humans by the LFNG gene.
Wingless-type MMTV integration site family, member 6, also known as WNT6, is a human gene.
This article is about the role of Fibroblast Growth Factor Signaling in Mesoderm Formation.

Mesoderm posterior protein 2 (MESP2), also known as class C basic helix-loop-helix protein 6 (bHLHc6), is a protein that in humans is encoded by the MESP2 gene.
Segmentation is the physical characteristic by which the human body is divided into repeating subunits called segments arranged along a longitudinal axis. In humans, the segmentation characteristic observed in the nervous system is of biological and evolutionary significance. Segmentation is a crucial developmental process involved in the patterning and segregation of groups of cells with different features, generating regional properties for such cell groups and organizing them both within the tissues as well as along the embryonic axis.

(HES7) or bHLHb37 is protein coding mammalian gene found on chromosome 17 in humans. HES7 is a member of the Hairy and Enhancer of Split families of Basic helix-loop-helix proteins. The gene product is a transcription factor and is expressed cyclically in the presomitic mesoderm as part of the Notch signalling pathway. HES7 is involved in the segmentation of somites from the presomitic mesoderm in vertebrates. The HES7 gene is self-regulated by a negative feedback loop in which the gene product can bind to its own promoter. This causes the gene to be expressed in an oscillatory manner. The HES7 protein also represses expression of Lunatic Fringe (LFNG) thereby both directly and indirectly regulating the Notch signalling pathway. Mutations in HES7 can result in deformities of the spine, ribs and heart. Spondylocostal dysostosis is a common disease caused by mutations in the HES7 gene. The inheritance pattern of Spondylocostal dysostosis is autosomal recessive.
Kim Dale FRSB, born in Cheltenham, England on 6 January 1971, is a research scientist based in the Cell and Developmental unit of School of Life Sciences at the University of Dundee, as well as being the Associate Dean International for UoD and the University Academic Regional Lead for ASEAN.
References
- ↑ Baker, R. E.; Schnell, S.; Maini, P. K. (2006). "A clock and wavefront mechanism for somite formation". Developmental Biology. 293 (1): 116–126. doi: 10.1016/j.ydbio.2006.01.018 . PMID 16546158.
- ↑ Goldbeter, A.; Pourquié, O. (2008). "Modeling the segmentation clock as a network of coupled oscillations in the Notch, Wnt and FGF signaling pathways". Journal of Theoretical Biology. 252 (3): 574–585. doi:10.1016/j.jtbi.2008.01.006. PMID 18308339.
- ↑ Gilbert, S.F. (2010). Developmental Biology (9th ed.). Sinauer Associates, Inc. pp. 413–415. ISBN 978-0-87893-384-6.
- ↑ Krol, A. J.; Roellig, D.; Dequéant, M. -L.; Tassy, O.; Glynn, E.; Hattem, G.; Mushegian, A.; Oates, A. C.; Pourquié, O. (2011). "Evolutionary plasticity of segmentation clock networks". Development. 138 (13): 2783–2792. doi:10.1242/dev.063834. PMC 3109603 . PMID 21652651.
- ↑ Gomez, C. L.; Özbudak, E. R. M.; Wunderlich, J.; Baumann, D.; Lewis, J.; Pourquié, O. (2008). "Control of segment number in vertebrate embryos". Nature. 454 (7202): 335–339. Bibcode:2008Natur.454..335G. doi:10.1038/nature07020. PMID 18563087. S2CID 4373389.
- ↑ Lewis, J.; Jiang, Y. J.; Aerne, B. L.; Smithers, L.; Haddon, C.; Ish-Horowicz, D. (2000). "Notch signalling and the synchronization of the somite segmentation clock". Nature. 408 (6811): 475–479. Bibcode:2000Natur.408..475J. doi:10.1038/35044091. PMID 11100729. S2CID 1182831.
- ↑ Resende, T. P.; Ferreira, M.; Teillet, M. -A.; Tavares, A. T.; Andrade, R. P.; Palmeirim, I. (2010). "Sonic hedgehog in temporal control of somite formation". Proceedings of the National Academy of Sciences. 107 (29): 12907–12912. Bibcode:2010PNAS..10712907R. doi: 10.1073/pnas.1000979107 . PMC 2919945 . PMID 20615943.