Related Research Articles

The striatum, or corpus striatum, is a nucleus in the subcortical basal ganglia of the forebrain. The striatum is a critical component of the motor and reward systems; receives glutamatergic and dopaminergic inputs from different sources; and serves as the primary input to the rest of the basal ganglia.

The olfactory bulb is a neural structure of the vertebrate forebrain involved in olfaction, the sense of smell. It sends olfactory information to be further processed in the amygdala, the orbitofrontal cortex (OFC) and the hippocampus where it plays a role in emotion, memory and learning. The bulb is divided into two distinct structures: the main olfactory bulb and the accessory olfactory bulb. The main olfactory bulb connects to the amygdala via the piriform cortex of the primary olfactory cortex and directly projects from the main olfactory bulb to specific amygdala areas. The accessory olfactory bulb resides on the dorsal-posterior region of the main olfactory bulb and forms a parallel pathway. Destruction of the olfactory bulb results in ipsilateral anosmia, while irritative lesions of the uncus can result in olfactory and gustatory hallucinations.

The claustrum is a thin, bilateral structure that connects to cortical and subcortical regions of the brain. It is located between the insula laterally and the putamen medially, separated by the extreme and external capsules respectively. The blood supply to the claustrum is fulfilled via the middle cerebral artery. It is considered to be the most densely connected structure in the brain, allowing for integration of various cortical inputs into one experience rather than singular events. The claustrum is difficult to study given the limited number of individuals with claustral lesions and the poor resolution of neuroimaging.
The nigrostriatal pathway is a bilateral dopaminergic pathway in the brain that connects the substantia nigra pars compacta (SNc) in the midbrain with the dorsal striatum in the forebrain. It is one of the four major dopamine pathways in the brain, and is critical in the production of movement as part of a system called the basal ganglia motor loop. Dopaminergic neurons of this pathway release dopamine from axon terminals that synapse onto GABAergic medium spiny neurons (MSNs), also known as spiny projection neurons (SPNs), located in the striatum.

A place cell is a kind of pyramidal neuron within the hippocampus that becomes active when an animal enters a particular place in its environment, which is known as the place field. Place cells are thought, collectively, to act as a cognitive representation of a specific location in space, known as a cognitive map. Place cells work with other types of neurons in the hippocampus and surrounding regions to perform this kind of spatial processing. They have been found in a variety of animals, including rodents, bats, monkeys and humans.

The periaqueductal gray is a nucleus that plays a critical role in autonomic function, motivated behavior and behavioural responses to threatening stimuli. PAG is also the primary control center for descending pain modulation. It has enkephalin-producing cells that suppress pain.

The thalamic reticular nucleus is part of the ventral thalamus that forms a capsule around the thalamus laterally. However, recent evidence from mice and fish question this statement and define it as a dorsal thalamic structure. It is separated from the thalamus by the external medullary lamina. Reticular cells are GABAergic, and have discoid dendritic arbors in the plane of the nucleus.
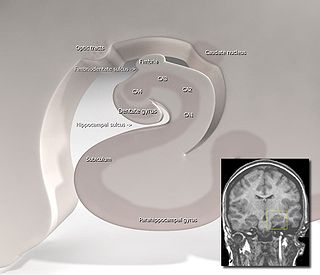
The subiculum is the most inferior component of the hippocampal formation. It lies between the entorhinal cortex and the CA1 subfield of the hippocampus proper.

Stellate cells are any neuron in the central nervous system that have a star-like shape formed by dendritic processes radiating from the cell body. Many Stellate cells are GABAergic and are located in the molecular layer of the cerebellum. Stellate cells are derived from dividing progenitors in the white matter of postnatal cerebellum. Dendritic trees can vary between neurons. There are two types of dendritic trees in the cerebral cortex, which include pyramidal cells, which are pyramid shaped and stellate cells which are star shaped. Dendrites can also aid neuron classification. Dendrites with spines are classified as spiny, those without spines are classified as aspinous. Stellate cells can be spiny or aspinous, while pyramidal cells are always spiny. Most common stellate cells are the inhibitory interneurons found within the upper half of the molecular layer in the cerebellum. Cerebellar stellate cells synapse onto the dendritic arbors of Purkinje cells and send inhibitory signals. Stellate neurons are sometimes found in other locations in the central nervous system; cortical spiny stellate cells are found in layer IVC of the V1 region in the visual cortex. In the somatosensory barrel cortex of mice and rats, glutamatergic (excitatory) spiny stellate cells are organized in the barrels of layer 4. They receive excitatory synaptic fibres from the thalamus and process feed forward excitation to 2/3 layer of V1 visual cortex to pyramidal cells. Cortical spiny stellate cells have a 'regular' firing pattern. Stellate cells are chromophobes, that is cells that does not stain readily, and thus appears relatively pale under the microscope.

The stria terminalis is a structure in the brain consisting of a band of fibers running along the lateral margin of the ventricular surface of the thalamus. Serving as a major output pathway of the amygdala, the stria terminalis runs from its centromedial division to the ventromedial nucleus of the hypothalamus.

Area 27 of Brodmann-1909 is a cytoarchitecturally defined cortical area that is a rostral part of the parahippocampal gyrus of the guenon (Brodmann-1909). It is commonly regarded as a synonym of presubiculum (Crosby-62).
The zona incerta is a horizontally elongated region of gray matter in the subthalamus below the thalamus. Its connections project extensively over the brain from the cerebral cortex down into the spinal cord.

The medial dorsal nucleus is a large nucleus in the thalamus.

The retrosplenial cortex (RSC) is a cortical area in the brain, located posteriorly and comprising Brodmann areas 29 and 30. The region's name refers to its anatomical location immediately behind the splenium of the corpus callosum in primates, although in rodents it is located more towards the brain surface and is relatively larger. Its function is currently not well understood, but its location close to visual areas and also to the hippocampal spatial/memory system suggest it may have a role in mediating between perceptual and memory functions.

Retrograde tracing is a research method used in neuroscience to trace neural connections from their point of termination to their source. Retrograde tracing techniques allow for detailed assessment of neuronal connections between a target population of neurons and their inputs throughout the nervous system. These techniques allow the "mapping" of connections between neurons in a particular structure and the target neurons in the brain. The opposite technique is anterograde tracing, which is used to trace neural connections from their source to their point of termination. Both the anterograde and retrograde tracing techniques are based on the visualization of axonal transport.
The parabrachial nuclei, also known as the parabrachial complex, are a group of nuclei in the dorsolateral pons that surrounds the superior cerebellar peduncle as it enters the brainstem from the cerebellum. They are named from the Latin term for the superior cerebellar peduncle, the brachium conjunctivum. In the human brain, the expansion of the superior cerebellar peduncle expands the parabrachial nuclei, which form a thin strip of grey matter over most of the peduncle. The parabrachial nuclei are typically divided along the lines suggested by Baxter and Olszewski in humans, into a medial parabrachial nucleus and lateral parabrachial nucleus. These have in turn been subdivided into a dozen subnuclei: the superior, dorsal, ventral, internal, external and extreme lateral subnuclei; the lateral crescent and subparabrachial nucleus along the ventrolateral margin of the lateral parabrachial complex; and the medial and external medial subnuclei
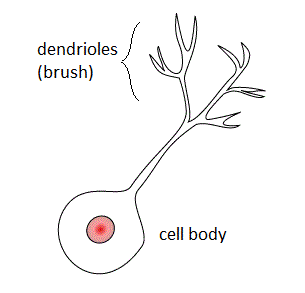
Unipolar brush cells (UBCs) are a class of excitatory glutamatergic interneuron found in the granular layer of the cerebellar cortex and also in the granule cell domain of the cochlear nucleus.
James B. Ranck Jr. is a distinguished professor of Physiology at the SUNY Downstate Medical Center. His research involves recording from single neurons in living animals for behavioral studies. He discovered head-direction cells in 1984.
The dorsal tegmental nucleus (DTN), also known as dorsal tegmental nucleus of Gudden (DTg), is a group of neurons located in the brain stem, which are involved in spatial navigation and orientation.
Laura Busse is a German neuroscientist and professor of Systemic Neuroscience within the Division of Neurobiology at the Ludwig Maximilian University of Munich. Busse's lab studies context-dependent visual processing in mouse models by performing large scale in vivo electrophysiological recordings in the thalamic and cortical circuits of awake and behaving mice.
References
- ↑ Taube, J. S.; Muller, R. U.; Ranck, J. B. (1990-02-01). "Head-direction cells recorded from the postsubiculum in freely moving rats. I. Description and quantitative analysis". The Journal of Neuroscience. 10 (2): 420–435. doi:10.1523/JNEUROSCI.10-02-00420.1990. ISSN 0270-6474. PMC 6570151 . PMID 2303851.
- ↑ Robertson, R. G.; Rolls, E. T.; Georges-François, P.; Panzeri, S. (1999-01-01). "Head direction cells in the primate pre-subiculum". Hippocampus. 9 (3): 206–219. doi:10.1002/(SICI)1098-1063(1999)9:3<206::AID-HIPO2>3.0.CO;2-H. ISSN 1050-9631. PMID 10401637.
- ↑ Yoder, Ryan M.; Taube, Jeffrey S. (2009-01-28). "Head direction cell activity in mice: robust directional signal depends on intact otolith organs". The Journal of Neuroscience. 29 (4): 1061–1076. doi:10.1523/JNEUROSCI.1679-08.2009. ISSN 1529-2401. PMC 2768409 . PMID 19176815.
- ↑ Muir, Gary M.; Brown, Joel E.; Carey, John P.; Hirvonen, Timo P.; Della Santina, Charles C.; Minor, Lloyd B.; Taube, Jeffrey S. (2009-11-18). "Disruption of the head direction cell signal after occlusion of the semicircular canals in the freely moving chinchilla". The Journal of Neuroscience. 29 (46): 14521–14533. doi:10.1523/JNEUROSCI.3450-09.2009. ISSN 1529-2401. PMC 2821030 . PMID 19923286.
- ↑ Rubin, Alon; Yartsev, Michael M.; Ulanovsky, Nachum (2014-01-15). "Encoding of head direction by hippocampal place cells in bats". The Journal of Neuroscience. 34 (3): 1067–1080. doi:10.1523/JNEUROSCI.5393-12.2014. ISSN 1529-2401. PMC 6608343 . PMID 24431464.
- ↑ Seelig, Johannes D.; Jayaraman, Vivek (2015-05-14). "Neural dynamics for landmark orientation and angular path integration". Nature. 521 (7551): 186–191. Bibcode:2015Natur.521..186S. doi:10.1038/nature14446. ISSN 1476-4687. PMC 4704792 . PMID 25971509.
- ↑ Taube, JS (2007). "The head direction signal: Origins and sensory-motor integration". Annu. Rev. Neurosci. 30: 181–207. doi:10.1146/annurev.neuro.29.051605.112854. PMID 17341158.
- ↑ Chen, L. L.; Lin, L. H.; Green, E. J.; Barnes, C. A.; McNaughton, B. L. (1994-01-01). "Head-direction cells in the rat posterior cortex. I. Anatomical distribution and behavioral modulation". Experimental Brain Research. 101 (1): 8–23. doi:10.1007/bf00243212. ISSN 0014-4819. PMID 7843305. S2CID 25125371.
- ↑ Giocomo, Lisa M.; Stensola, Tor; Bonnevie, Tora; Van Cauter, Tiffany; Moser, May-Britt; Moser, Edvard I. (2014-02-03). "Topography of head direction cells in medial entorhinal cortex". Current Biology. 24 (3): 252–262. doi: 10.1016/j.cub.2013.12.002 . ISSN 1879-0445. PMID 24440398.
- ↑ Taube, J. S. (1995-01-01). "Head direction cells recorded in the anterior thalamic nuclei of freely moving rats". The Journal of Neuroscience. 15 (1 Pt 1): 70–86. doi:10.1523/JNEUROSCI.15-01-00070.1995. ISSN 0270-6474. PMC 6578288 . PMID 7823153.
- ↑ Mizumori, S. J.; Williams, J. D. (1993-09-01). "Directionally selective mnemonic properties of neurons in the lateral dorsal nucleus of the thalamus of rats". The Journal of Neuroscience. 13 (9): 4015–4028. doi:10.1523/JNEUROSCI.13-09-04015.1993. ISSN 0270-6474. PMC 6576470 . PMID 8366357.
- ↑ Stackman, R. W.; Taube, J. S. (1998-11-01). "Firing properties of rat lateral mammillary single units: head direction, head pitch, and angular head velocity". The Journal of Neuroscience. 18 (21): 9020–9037. doi:10.1523/JNEUROSCI.18-21-09020.1998. ISSN 0270-6474. PMC 1550347 . PMID 9787007.
- ↑ Yoder, Ryan M.; Taube, Jeffrey S. (2014-01-01). "The vestibular contribution to the head direction signal and navigation". Frontiers in Integrative Neuroscience. 8: 32. doi:10.3389/fnint.2014.00032. PMC 4001061 . PMID 24795578.
- ↑ Jacob, Pierre-Yves; Casali, Giulio; Spieser, Laure; Page, Hector; Overington, Dorothy; Jeffery, Kate (2016-12-19). "An independent, landmark-dominated head-direction signal in dysgranular retrosplenial cortex". Nature Neuroscience. 20 (2): 173–175. doi:10.1038/nn.4465. ISSN 1546-1726. PMC 5274535 . PMID 27991898.
- ↑ O'Keefe, J.; Dostrovsky, J. (1971-11-01). "The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat". Brain Research. 34 (1): 171–175. doi:10.1016/0006-8993(71)90358-1. ISSN 0006-8993. PMID 5124915.
- ↑ Golob, E. J.; Taube, J. S. (1999-08-15). "Head direction cells in rats with hippocampal or overlying neocortical lesions: evidence for impaired angular path integration". The Journal of Neuroscience. 19 (16): 7198–7211. doi:10.1523/JNEUROSCI.19-16-07198.1999. ISSN 1529-2401. PMC 6782884 . PMID 10436073.
- ↑ Blair, H. T.; Sharp, P. E. (1996-08-01). "Visual and vestibular influences on head-direction cells in the anterior thalamus of the rat". Behavioral Neuroscience. 110 (4): 643–660. doi:10.1037/0735-7044.110.4.643. ISSN 0735-7044. PMID 8864258.
- ↑ Raudies, Florian; Brandon, Mark P.; Chapman, G. William; Hasselmo, Michael E. (2015-09-24). "Head direction is coded more strongly than movement direction in a population of entorhinal neurons". Brain Research. 1621: 355–367. doi:10.1016/j.brainres.2014.10.053. ISSN 1872-6240. PMC 4427560 . PMID 25451111.
- ↑ Blair, H. T.; Sharp, P. E. (1995-09-01). "Anticipatory head direction signals in anterior thalamus: evidence for a thalamocortical circuit that integrates angular head motion to compute head direction". The Journal of Neuroscience. 15 (9): 6260–6270. doi:10.1523/JNEUROSCI.15-09-06260.1995. ISSN 0270-6474. PMC 6577663 . PMID 7666208.
- ↑ Peyrache, A; Lacroix MM; Petersen PC; Buzsaki G (2015). "Internally organized mechanisms of the head direction sense". Nat. Neurosci. 18 (4): 569–575. doi:10.1038/nn.3968. PMC 4376557 . PMID 25730672.
- ↑ Yoder, Ryan M.; Taube, Jeffrey S. (2014-01-01). "The vestibular contribution to the head direction signal and navigation". Frontiers in Integrative Neuroscience. 8: 32. doi:10.3389/fnint.2014.00032. PMC 4001061 . PMID 24795578.
- ↑ Tryon, Valerie L.; Kim, Esther U.; Zafar, Talal J.; Unruh, April M.; Staley, Shelly R.; Calton, Jeffrey L. (2012-12-01). "Magnetic field polarity fails to influence the directional signal carried by the head direction cell network and the behavior of rats in a task requiring magnetic field orientation". Behavioral Neuroscience. 126 (6): 835–844. doi:10.1037/a0030248. ISSN 1939-0084. PMID 23025828.
- ↑ Zhang, K. (1996-03-15). "Representation of spatial orientation by the intrinsic dynamics of the head-direction cell ensemble: a theory". The Journal of Neuroscience. 16 (6): 2112–2126. doi:10.1523/JNEUROSCI.16-06-02112.1996. ISSN 0270-6474. PMC 6578512 . PMID 8604055.
- ↑ Seelig, JD; Jayaraman V (May 14, 2015). "Neural dynamics for landmark orientation and angular path integration". Nature. 521 (7551): 186–191. Bibcode:2015Natur.521..186S. doi:10.1038/nature14446. PMC 4704792 . PMID 25971509.
- ↑ Song, Pengcheng; Wang, Xiao-Jing (2005-01-26). "Angular Path Integration by Moving "Hill of Activity": A Spiking Neuron Model without Recurrent Excitation of the Head-Direction System". Journal of Neuroscience. 25 (4): 1002–1014. doi:10.1523/jneurosci.4172-04.2005. PMC 6725619 . PMID 15673682.
- ↑ Ranck Jr, J. B. "Head direction cells in the deep cell layer of dorsal presubiculum in freely moving rats." Soc Neurosci Abstr. Vol. 10. No. 176.12. 1984.
- ↑ Taube, JS; Muller RU; Ranck JB Jr. (1 February 1990). "Head-direction cells recorded from the postsubiculum in freely moving rats. I. Description and quantitative analysis". J. Neurosci. 10 (2): 420–435. doi: 10.1523/JNEUROSCI.10-02-00420.1990 . PMID 2303851.
- ↑ Taube, JS; Muller, RU; Ranck, JB (February 1990). "Head-direction cells recorded from the postsubiculum in freely moving rats. II. Effects of environmental manipulations". J. Neurosci. 10 (2): 436–447. doi:10.1523/JNEUROSCI.10-02-00436.1990. PMC 6570161 . PMID 2303852.
- ↑ Mizumori, SJ; Williams JD (September 1, 1993). "Directionally selective mnemonic properties of neurons in the lateral dorsal nucleus of the thalamus of rats". J. Neurosci. 13 (9): 4015–4028. doi: 10.1523/JNEUROSCI.13-09-04015.1993 . PMID 8366357.
- ↑ Taube, JS (January 1, 1995). "Head direction cells recorded in the anterior thalamic nuclei of freely moving rats". J. Neurosci. 15 (1): 70–86. doi: 10.1523/JNEUROSCI.15-01-00070.1995 . PMID 7823153.
- ↑ Chen, LL; Lin LH; Green EJ; Barnes CA; McNaughton BL (1994). "Head-direction cells in the rat posterior cortex. I. Anatomical distribution and behavioral modulation". Exp. Brain Res. 101 (1): 8–23. doi:10.1007/BF00243212. PMID 7843305. S2CID 25125371.
- ↑ Zhang, K (March 15, 1996). "Representation of spatial orientation by the intrinsic dynamics of the head-direction cell ensemble: a theory". J. Neurosci. 16 (6): 2112–2126. doi: 10.1523/JNEUROSCI.16-06-02112.1996 . PMID 8604055.
- ↑ Robertson, RG; Rolls ET; Georges-François P; Panzeri S (1999). "Head direction cells in the primate pre-subiculum". Hippocampus. 9 (3): 206–19. doi:10.1002/(sici)1098-1063(1999)9:3<206::aid-hipo2>3.0.co;2-h. PMID 10401637.
- ↑ Finkelstein, A; Derdikman D; Rubin A; Foerster JN; Las L; Ulanovsky N (January 8, 2015). "Three-dimensional head-direction coding in the bat brain". Nature. 517 (7533): 159–164. Bibcode:2015Natur.517..159F. doi:10.1038/nature14031. PMID 25470055. S2CID 4457477.