Paraoxonase 1 (PON1) also known as Serum paraoxonase and arylesterase 1, A esterase, homocysteine thiolactonase or serum aryldialkylphosphatase 1, is an enzyme that in humans is encoded by the PON1gene.[5] Paraoxonase 1 has esterase and more specifically paraoxonase activity.[6]PON1 is the first discovered member of a multigene family also containing PON2 and PON3, the genes for which are located adjacent to each other on chromosome 7. PON1 on HDL (different from soluble PON1) is responsible for significant atheroprotection rendered by the HDL.[7]
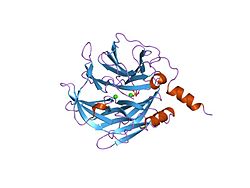
Human PON1 is a glycoprotein composed of 354 amino acids and has a molecular weight of 43,000 daltons which associates with high-density lipoprotein (HDL) in the circulation. Serum PON1 is secreted mainly by the liver, although local synthesis occurs in several tissues and PON1 protein is found in almost all tissues. X-ray crystallography has revealed the structure of PON1 to be a 6 bladed propeller with a unique lid structure covering the active site passage which allows association with HDL.[8][9][10]
PON1 is a highly promiscuous enzyme capable of hydrolysing a wide variety of substrates, such as lactones,[15] including thiolactones and pharmaceutical agents such as statins. PON1 substrates also include glucuronide drugs, arylesters, cyclic carbonates, organophosphorus pesticides and nerve gases such as sarin, soman and VX, oestrogen esters and lipid peroxides (oxidized lipids). Oxidized polyunsaturated fatty acids (notably in oxidized low-density lipoprotein) form lactone-like structures that are PON substrates.[16]
Genetics
PON1 in humans is encoded by the PON1 gene, which is located on the long arm of chromosome 7.[17] PON1 activity can vary by over 40 fold between individuals. The biggest effect on PON1 activity levels is through PON1 genetic polymorphisms.[9] Many nutritional, life-style and pharmaceutical modulators of PON1 are also known.[18][19]
The coding region PON1-Q192R polymorphism determines a substrate dependent effect on activity. Many organophosphates used in pesticides, such as paraoxon, are hydrolysed faster by the PON1-R allozyme. Other substrates such as diazoxon, lipid-peroxides and sarin are hydrolysed more rapidly by the PON1-Q allozyme.[9]
Both the coding region PON1-L55M and the promoter region PON1-T-108C polymorphisms are associated with different serum concentrations and therefore activities. The 55L allele results in significantly higher PON1 mRNA and serum protein levels and therefore activity compared to the 55M allele.[18][19] The -108C allele has greater promoter activity than the -108T allele which results in different serum activities.[18][19]
The distribution of the PON1 polymorphisms varies with ethnicity. The frequency of the PON1-192R allele increases the further from Europe a population originates, the frequency in Caucasians of 15-30% increases to 70-90% in Far Eastern Oriental and Sub-Saharan African populations.[20] In the southern US, African-Americans are five times more likely to be RR than Caucasians.[21] In contrast, the PON1-55M allele is much less frequent in Oriental and black African populations compared to Caucasians and are extremely rare or absent in some populations e.g. Thais. These ethnic differences in SNP distribution can lead to large activity differences between populations.[20]
Clinical significance
PON1 was first discovered through its ability to hydrolyse and therefore detoxify organophosphorus compounds which are widely used as pesticides and nerve gases. PON1 protects humans from the acute and chronic harmful effects of these compounds.[22][23] Low PON1 activity found in children may increase their susceptibility to organophosphates. Because PON1-Q is more protective against sarin than PON1-R, the PON1-R allele and low PON1-Q activity levels are associated with Gulf War Illness.[24]
The greatest research interest has been the role of PON1 in atherosclerosis, where, because of its ability to remove harmful oxidised-lipids, PON1 protects against the development of atherosclerosis.[25]
PON1 also protects against bacterial infection by destroying the bacterial signalling molecules that cause gram negative bacteria to invade human tissue and form colonies, thus PON1 contributes to the bodies innate immunity.[26]
Recently it has been suggested that PON1 has a role in healthy aging, however, the mechanism is currently unknown.[27]
PON1 activity is low in infants compared to adults. A study of Mexican-American children showed that PON1 activity increased 3.5 times between birth and age seven.[28]
An association between PON1 gene polymorphism and susceptibility to Parkinson's disease was not found in a Chinese population.[29]
↑"Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
↑"Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
↑Primo-Parmo SL, Sorenson RC, Teiber J, La Du BN (May 1996). "The human serum paraoxonase/arylesterase gene (PON1) is one member of a multigene family". Genomics. 33 (3): 498–507. doi:10.1006/geno.1996.0225. PMID8661009.
↑Harel M, Aharoni A, Gaidukov L, Brumshtein B, Khersonsky O, Meged R, etal. (May 2004). "Structure and evolution of the serum paraoxonase family of detoxifying and anti-atherosclerotic enzymes". Nature Structural & Molecular Biology. 11 (5): 412–9. doi:10.1038/nsmb767. PMID15098021. S2CID52874893.
↑Deakin SP, James RW (Nov 2004). "Genetic and environmental factors modulating serum concentrations and activities of the antioxidant enzyme paraoxonase-1". Clinical Science. 107 (5): 435–47. doi:10.1042/CS20040187. PMID15265000. S2CID18754293.
↑Costa LG, Cole TB, Vitalone A, Furlong CE (Feb 2005). "Measurement of paraoxonase (PON1) status as a potential biomarker of susceptibility to organophosphate toxicity". Clinica Chimica Acta; International Journal of Clinical Chemistry. 352 (1–2): 37–47. doi:10.1016/j.cccn.2004.09.019. PMID15653099.
↑Mackness M, Mackness B (Nov 2004). "Paraoxonase 1 and atherosclerosis: is the gene or the protein more important?". Free Radical Biology & Medicine. 37 (9): 1317–23. doi:10.1016/j.freeradbiomed.2004.07.034. PMID15454272.
↑Khateeb J, Gantman A, Kreitenberg AJ, Aviram M, Fuhrman B (Jan 2010). "Paraoxonase 1 (PON1) expression in hepatocytes is upregulated by pomegranate polyphenols: a role for PPAR-gamma pathway". Atherosclerosis. 208 (1): 119–25. doi:10.1016/j.atherosclerosis.2009.08.051. PMID19783251.
↑Khersonsky O, Tawfik DS (Apr 2005). "Structure-reactivity studies of serum paraoxonase PON1 suggest that its native activity is lactonase". Biochemistry. 44 (16): 6371–82. doi:10.1021/bi047440d. PMID15835926.
12La Du BN (1992). "Human serum paraoxonase/arylesterase". In Kalow W (ed.). Pharmacogenetics of Drug Metabolism. New York: Pergamon Press. pp.51–91.
↑McDaniel CY, Dail MB, Wills RW, Chambers HW, Chambers JE (Dec 2014). "Paraoxonase 1 polymorphisms within a Mississippi USA population as possible biomarkers of enzyme activities associated with disease susceptibility". Biochemical Genetics. 52 (11–12): 509–23. doi:10.1007/s10528-014-9663-8. PMID25027835. S2CID16649798.
↑Mackness M, Mackness B (2014). "Current aspects of paraoxonase-1 research". In Komoda T (ed.). The HDL handbook: biological functions and clinical implications (Seconded.). Amsterdam: Academic Press. ISBN978-0-12-407867-3.
↑Costa LG, Cole TB, Jarvik GP, Furlong CE (2003). "Functional genomic of the paraoxonase (PON1) polymorphisms: effects on pesticide sensitivity, cardiovascular disease, and drug metabolism". Annual Review of Medicine. 54: 371–92. doi:10.1146/annurev.med.54.101601.152421. PMID12525679.
Costa LG, Cole TB, Furlong CE (2003). "Polymorphisms of paraoxonase (PON1) and their significance in clinical toxicology of organophosphates". Journal of Toxicology. Clinical Toxicology. 41 (1): 37–45. doi:10.1081/CLT-120018269. PMID12645966. S2CID46233526.
Furlong CE, Cole TB, Jarvik GP, Pettan-Brewer C, Geiss GK, Richter RJ, etal. (Aug 2005). "Role of paraoxonase (PON1) status in pesticide sensitivity: genetic and temporal determinants". Neurotoxicology. 26 (4): 651–9. doi:10.1016/j.neuro.2004.08.002. PMID16112327.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.