Related Research Articles

The inner ear is the innermost part of the vertebrate ear. In vertebrates, the inner ear is mainly responsible for sound detection and balance. In mammals, it consists of the bony labyrinth, a hollow cavity in the temporal bone of the skull with a system of passages comprising two main functional parts:

Pitch is a perceptual property that allows sounds to be ordered on a frequency-related scale, or more commonly, pitch is the quality that makes it possible to judge sounds as "higher" and "lower" in the sense associated with musical melodies. Pitch is a major auditory attribute of musical tones, along with duration, loudness, and timbre.

The cochlea is the part of the inner ear involved in hearing. It is a spiral-shaped cavity in the bony labyrinth, in humans making 2.75 turns around its axis, the modiolus. A core component of the cochlea is the organ of Corti, the sensory organ of hearing, which is distributed along the partition separating the fluid chambers in the coiled tapered tube of the cochlea.
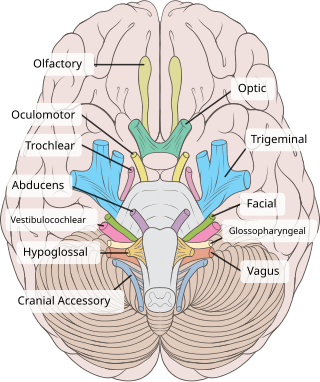
The vestibulocochlear nerve or auditory vestibular nerve, also known as the eighth cranial nerve, cranial nerve VIII, or simply CN VIII, is a cranial nerve that transmits sound and equilibrium (balance) information from the inner ear to the brain. Through olivocochlear fibers, it also transmits motor and modulatory information from the superior olivary complex in the brainstem to the cochlea.

The basilar membrane is a stiff structural element within the cochlea of the inner ear which separates two liquid-filled tubes that run along the coil of the cochlea, the scala media and the scala tympani. The basilar membrane moves up and down in response to incoming sound waves, which are converted to traveling waves on the basilar membrane.
Place theory is a theory of hearing that states that our perception of sound depends on where each component frequency produces vibrations along the basilar membrane. By this theory, the pitch of a sound, such as a human voice or a musical tone, is determined by the places where the membrane vibrates, based on frequencies corresponding to the tonotopic organization of the primary auditory neurons.
Stimulus modality, also called sensory modality, is one aspect of a stimulus or what is perceived after a stimulus. For example, the temperature modality is registered after heat or cold stimulate a receptor. Some sensory modalities include: light, sound, temperature, taste, pressure, and smell. The type and location of the sensory receptor activated by the stimulus plays the primary role in coding the sensation. All sensory modalities work together to heighten stimuli sensation when necessary.

The auditory system is the sensory system for the sense of hearing. It includes both the sensory organs and the auditory parts of the sensory system.

The acoustic reflex is an involuntary muscle contraction that occurs in the middle ear in response to loud sound stimuli or when the person starts to vocalize.
Sensorineural hearing loss (SNHL) is a type of hearing loss in which the root cause lies in the inner ear, sensory organ, or the vestibulocochlear nerve. SNHL accounts for about 90% of reported hearing loss. SNHL is usually permanent and can be mild, moderate, severe, profound, or total. Various other descriptors can be used depending on the shape of the audiogram, such as high frequency, low frequency, U-shaped, notched, peaked, or flat.
In physiology, tonotopy is the spatial arrangement of where sounds of different frequency are processed in the brain. Tones close to each other in terms of frequency are represented in topologically neighbouring regions in the brain. Tonotopic maps are a particular case of topographic organization, similar to retinotopy in the visual system.

Volley theory states that groups of neurons of the auditory system respond to a sound by firing action potentials slightly out of phase with one another so that when combined, a greater frequency of sound can be encoded and sent to the brain to be analyzed. The theory was proposed by Ernest Wever and Charles Bray in 1930 as a supplement to the frequency theory of hearing. It was later discovered that this only occurs in response to sounds that are about 500 Hz to 5000 Hz.

The cochlear nucleus (CN) or cochlear nuclear complex comprises two cranial nerve nuclei in the human brainstem, the ventral cochlear nucleus (VCN) and the dorsal cochlear nucleus (DCN). The ventral cochlear nucleus is unlayered whereas the dorsal cochlear nucleus is layered. Auditory nerve fibers, fibers that travel through the auditory nerve carry information from the inner ear, the cochlea, on the same side of the head, to the nerve root in the ventral cochlear nucleus. At the nerve root the fibers branch to innervate the ventral cochlear nucleus and the deep layer of the dorsal cochlear nucleus. All acoustic information thus enters the brain through the cochlear nuclei, where the processing of acoustic information begins. The outputs from the cochlear nuclei are received in higher regions of the auditory brainstem.

The interaural time difference when concerning humans or animals, is the difference in arrival time of a sound between two ears. It is important in the localization of sounds, as it provides a cue to the direction or angle of the sound source from the head. If a signal arrives at the head from one side, the signal has further to travel to reach the far ear than the near ear. This pathlength difference results in a time difference between the sound's arrivals at the ears, which is detected and aids the process of identifying the direction of sound source.
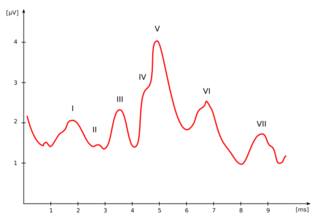
The auditory brainstem response (ABR), also called brainstem evoked response audiometry (BERA) or brainstem auditory evoked potentials (BAEPs) or brainstem auditory evoked responses (BAERs) is an auditory evoked potential extracted from ongoing electrical activity in the brain and recorded via electrodes placed on the scalp. The measured recording is a series of six to seven vertex positive waves of which I through V are evaluated. These waves, labeled with Roman numerals in Jewett and Williston convention, occur in the first 10 milliseconds after onset of an auditory stimulus. The ABR is considered an exogenous response because it is dependent upon external factors.
Computational auditory scene analysis (CASA) is the study of auditory scene analysis by computational means. In essence, CASA systems are "machine listening" systems that aim to separate mixtures of sound sources in the same way that human listeners do. CASA differs from the field of blind signal separation in that it is based on the mechanisms of the human auditory system, and thus uses no more than two microphone recordings of an acoustic environment. It is related to the cocktail party problem.
In audio signal processing, auditory masking occurs when the perception of one sound is affected by the presence of another sound.
Hearing, or auditory perception, is the ability to perceive sounds through an organ, such as an ear, by detecting vibrations as periodic changes in the pressure of a surrounding medium. The academic field concerned with hearing is auditory science.
Electrocochleography is a technique of recording electrical potentials generated in the inner ear and auditory nerve in response to sound stimulation, using an electrode placed in the ear canal or tympanic membrane. The test is performed by an otologist or audiologist with specialized training, and is used for detection of elevated inner ear pressure or for the testing and monitoring of inner ear and auditory nerve function during surgery.
Temporal envelope (ENV) and temporal fine structure (TFS) are changes in the amplitude and frequency of sound perceived by humans over time. These temporal changes are responsible for several aspects of auditory perception, including loudness, pitch and timbre perception and spatial hearing.
References
- ↑ James Beament (2001). How We Hear Music . The Boydell Press. ISBN 0-85115-813-7.
- ↑ Nelson Y. S. Kiang (1969). Discharge Patterns of Single Auditory Fibers. MIT Research Monograph 35.
- ↑ J. J. Rose; J. Hind; D. Anderson & J. Brugge (1967). "Response of Auditory Fibers in the Squirrel Monkey". J. Neurophysiol. 30 (4): 769–793. doi:10.1152/jn.1967.30.4.769. PMID 4962851.
- ↑ Alain de Cheveigné (2005). "Pitch Perception Models". In Christopher J. Plack; Andrew J. Oxenham; Richard R. Fay; Arthur N. Popper (eds.). Pitch. Birkhäuser. ISBN 0-387-23472-1.
- ↑ Shackleton, Trevor M.; Carlyon, Robert (1994). "The role of resolved and unresolved harmonics in pitch perception and frequency modulation discrimination". The Journal of the Acoustical Society of America. 95 (6): 3529. doi: 10.1121/1.409970 . Retrieved 22 November 2016.
- ↑ Fearn R, Carter P, Wolfe J (1999). "The perception of pitch by users of cochlear implants: possible significance for rate and place theories of pitch". Acoustics Australia. 27 (2): 41–43.