Homininae, also called "African hominids" or "African apes", is a subfamily of Hominidae. It includes two tribes, with their extant as well as extinct species: 1) the tribe Hominini ―and 2) the tribe Gorillini (gorillas). Alternatively, the genus Pan is sometimes considered to belong to its own third tribe, Panini. Homininae comprises all hominids that arose after orangutans split from the line of great apes. The Homininae cladogram has three main branches, which lead to gorillas and to humans and chimpanzees. There are two living species of Panina and two living species of gorillas, but only one extant human species. Traces of extinct Homo species, including Homo floresiensis have been found with dates as recent as 40,000 years ago. Organisms in this subfamily are described as hominine or hominines.
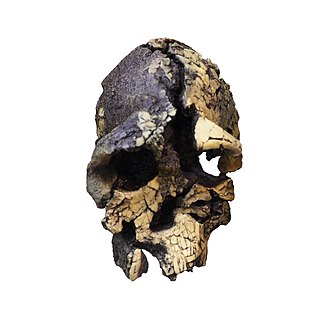
Kenyanthropus is a genus of extinct hominin identified from the Lomekwi site by Lake Turkana, Kenya, dated to 3.3 to 3.2 million years ago during the Middle Pliocene. It contains one species, K. platyops, but may also include the 2 million year old Homo rudolfensis, or K. rudolfensis. Before its naming in 2001, Australopithecus afarensis was widely regarded as the only australopithecine to exist during the Middle Pliocene, but Kenyanthropus evinces a greater diversity than once acknowledged. Kenyanthropus is most recognisable by an unusually flat face and small teeth for such an early hominin, with values on the extremes or beyond the range of variation for australopithecines in regard to these features. Multiple australopithecine species may have coexisted by foraging for different food items, which may be reason why these apes anatomically differ in features related to chewing.

Orrorin is an extinct genus of primate within Homininae from the Miocene Lukeino Formation and Pliocene Mabaget Formation, both of Kenya.

Australopithecus afarensis is an extinct species of australopithecine which lived from about 3.9–2.9 million years ago (mya) in the Pliocene of East Africa. The first fossils were discovered in the 1930s, but major fossil finds would not take place until the 1970s. From 1972 to 1977, the International Afar Research Expedition—led by anthropologists Maurice Taieb, Donald Johanson and Yves Coppens—unearthed several hundreds of hominin specimens in Hadar, Ethiopia, the most significant being the exceedingly well-preserved skeleton AL 288-1 ("Lucy") and the site AL 333. Beginning in 1974, Mary Leakey led an expedition into Laetoli, Tanzania, and notably recovered fossil trackways. In 1978, the species was first described, but this was followed by arguments for splitting the wealth of specimens into different species given the wide range of variation which had been attributed to sexual dimorphism. A. afarensis probably descended from A. anamensis and is hypothesised to have given rise to Homo, though the latter is debated.
Paleoanthropology or paleo-anthropology is a branch of paleontology and anthropology which seeks to understand the early development of anatomically modern humans, a process known as hominization, through the reconstruction of evolutionary kinship lines within the family Hominidae, working from biological evidence and cultural evidence.

Sterkfontein is a set of limestone caves of special interest in paleoanthropology located in Gauteng province, about 40 kilometres (25 mi) northwest of Johannesburg, South Africa in the Muldersdrift area close to the town of Krugersdorp. The archaeological sites of Swartkrans and Kromdraai are in the same area. Sterkfontein is a South African National Heritage Site and was also declared a World Heritage Site in 2000. The area in which it is situated is known as the Cradle of Humankind. The Sterkfontein Caves are also home to numerous wild African species including Belonogaster petiolata, a wasp species of which there is a large nesting presence.

Australopithecus africanus is an extinct species of australopithecine which lived between about 3.3 and 2.1 million years ago in the Late Pliocene to Early Pleistocene of South Africa. The species has been recovered from Taung, Sterkfontein, Makapansgat, and Gladysvale. The first specimen, the Taung child, was described by anatomist Raymond Dart in 1924, and was the first early hominin found. However, its closer relations to humans than to other apes would not become widely accepted until the middle of the century because most had believed humans evolved outside of Africa. It is unclear how A. africanus relates to other hominins, being variously placed as ancestral to Homo and Paranthropus, to just Paranthropus, or to just P. robustus. The specimen "Little Foot" is the most completely preserved early hominin, with 90% of the skeleton intact, and the oldest South African australopith. However, it is controversially suggested that it and similar specimens be split off into "A. prometheus".
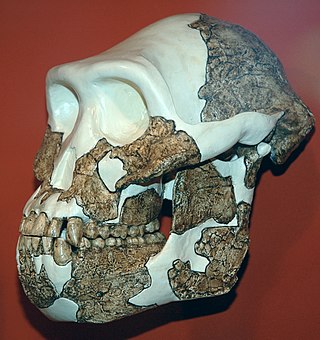
Australopithecus anamensis is a hominin species that lived approximately between 4.2 and 3.8 million years ago and is the oldest known Australopithecus species, living during the Plio-Pleistocene era.

Paranthropus aethiopicus is an extinct species of robust australopithecine from the Late Pliocene to Early Pleistocene of East Africa about 2.7–2.3 million years ago. However, it is much debated whether or not Paranthropus is an invalid grouping and is synonymous with Australopithecus, so the species is also often classified as Australopithecus aethiopicus. Whatever the case, it is considered to have been the ancestor of the much more robust P. boisei. It is debated if P. aethiopicus should be subsumed under P. boisei, and the terms P. boisei sensu lato and P. boisei sensu stricto can be used to respectively include and exclude P. aethiopicus from P. boisei.

Australopithecus garhi is a species of australopithecine from the Bouri Formation in the Afar Region of Ethiopia 2.6–2.5 million years ago (mya) during the Early Pleistocene. The first remains were described in 1999 based on several skeletal elements uncovered in the three years preceding. A. garhi was originally considered to have been a direct ancestor to Homo and the human line, but is now thought to have been an offshoot. Like other australopithecines, A. garhi had a brain volume of 450 cc (27 cu in); a jaw which jutted out (prognathism); relatively large molars and premolars; adaptations for both walking on two legs (bipedalism) and grasping while climbing (arboreality); and it is possible that, though unclear if, males were larger than females. One individual, presumed female based on size, may have been 140 cm tall.

Paranthropus robustus is a species of robust australopithecine from the Early and possibly Middle Pleistocene of the Cradle of Humankind, South Africa, about 2.27 to 0.87 million years ago. It has been identified in Kromdraai, Swartkrans, Sterkfontein, Gondolin, Cooper's, and Drimolen Caves. Discovered in 1938, it was among the first early hominins described, and became the type species for the genus Paranthropus. However, it has been argued by some that Paranthropus is an invalid grouping and synonymous with Australopithecus, so the species is also often classified as Australopithecus robustus.

Paranthropus boisei is a species of australopithecine from the Early Pleistocene of East Africa about 2.5 to 1.15 million years ago. The holotype specimen, OH 5, was discovered by palaeoanthropologist Mary Leakey in 1959 at Olduvai Gorge, Tanzania and described by her husband Louis a month later. It was originally placed into its own genus as "Zinjanthropus boisei", but is now relegated to Paranthropus along with other robust australopithecines. However, it is also argued that Paranthropus is an invalid grouping and synonymous with Australopithecus, so the species is also often classified as Australopithecus boisei.

The Hominini form a taxonomic tribe of the subfamily Homininae ("hominines"). Hominini includes the extant genera Homo (humans) and Pan and in standard usage excludes the genus Gorilla (gorillas).
Orthograde is a term derived from Greek ὀρθός, orthos + Latin gradi that describes a manner of walking which is upright, with the independent motion of limbs. Both New and Old World monkeys are primarily arboreal, and they have a tendency to walk with their limbs swinging in parallel to one another. This differs from the manner of walking demonstrated by the apes.

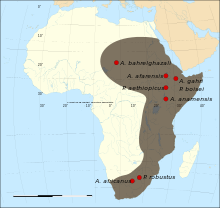
The australopithecines, formally Australopithecina or Hominina, are generally any species in the related genera of Australopithecus and Paranthropus. It may also include members of Kenyanthropus, Ardipithecus, and Praeanthropus. The term comes from a former classification as members of a distinct subfamily, the Australopithecinae. They are now classified within the Australopithecina subtribe of the Hominini tribe. All these related species are now sometimes collectively termed australopithecines, australopiths or homininians. They are the extinct, close relatives of modern humans and, together with the extant genus Homo, comprise the human clade. Members of the human clade, i.e. the Hominini after the split from the chimpanzees, are now called Hominina.
Australopithecus bahrelghazali is an extinct species of australopithecine discovered in 1995 at Koro Toro, Bahr el Gazel, Chad, existing around 3.5 million years ago in the Pliocene. It is the first and only australopithecine known from Central Africa, and demonstrates that this group was widely distributed across Africa as opposed to being restricted to East and southern Africa as previously thought. The validity of A. bahrelghazali has not been widely accepted, in favour of classifying the specimens as A. afarensis, a better known Pliocene australopithecine from East Africa, because of the anatomical similarity and the fact that A. bahrelghazali is known only from 3 partial jawbones and an isolated premolar. The specimens inhabited a lakeside grassland environment with sparse tree cover, possibly similar to the modern Okavango Delta, and similarly predominantly ate C4 savanna foods—such as grasses, sedges, storage organs, or rhizomes—and to a lesser degree also C3 forest foods—such as fruits, flowers, pods, or insects. However, the teeth seem ill-equipped to process C4 plants, so its true diet is unclear.
The Middle Awash is a paleoanthropological research area in the northwest corner of Gabi Rasu in the Afar Region along the Awash River in Ethiopia's Afar Depression. It is a unique natural laboratory for the study of human origins and evolution and a number of fossils of the earliest hominins, particularly of the Australopithecines, as well as some of the oldest known Olduwan stone artifacts, have been found at the site—all of late Miocene, the Pliocene, and the very early Pleistocene times, that is, about 5.6 million years ago (mya) to 2.5 mya. It is broadly thought that the divergence of the lines of the earliest humans (hominins) and of chimpanzees (hominids) was completed near the beginning of that time range, or sometime between seven and five mya. However, the larger community of scientists provide several estimates for periods of divergence that imply a greater range for this event, see CHLCA: human-chimpanzee split.

Post-canine megadontia is a relative enlargement of the molars and premolars compared to the size of the incisors and canines. This phenomenon is seen in some early hominid ancestors such as Paranthropus aethiopicus.

Australopithecus sediba is an extinct species of australopithecine recovered from Malapa Cave, Cradle of Humankind, South Africa. It is known from a partial juvenile skeleton, the holotype MH1, and a partial adult female skeleton, the paratype MH2. They date to about 1.98 million years ago in the Early Pleistocene, and coexisted with Paranthropus robustus and Homo ergaster / Homo erectus. Malapa is interpreted as having been a natural death trap, the base of a long vertical shaft which creatures could accidentally fall into. A. sediba was initially described as being a potential human ancestor, and perhaps the progenitor of Homo, but this is contested and it could also represent a late-surviving population or sister species of A. africanus which had earlier inhabited the area.
Australopithecus deyiremeda is an extinct species of australopithecine from Woranso–Mille, Afar Region, Ethiopia, about 3.5 to 3.3 million years ago during the Pliocene. Because it is known only from three partial jawbones, it is unclear if these specimens indeed represent a unique species or belong to the much better-known A. afarensis. A. deyiremeda is distinguished by its forward-facing cheek bones and small cheek teeth compared to those of other early hominins. It is unclear if a partial foot specimen exhibiting a dextrous big toe can be assigned to A. deyiremeda. A. deyiremeda lived in a mosaic environment featuring both open grasslands and lake- or riverside forests, and anthropologist Fred Spoor suggests it may have been involved in the Kenyan Lomekwi stone-tool industry typically assigned to Kenyanthropus. A. deyiremeda coexisted with A. afarensis, and they may have exhibited niche partitioning to avoid competing with each other for the same resources, such as by relying on different fallback foods during leaner times.