
A chordate is a deuterostomic animal belonging to the phylum Chordata. All chordates possess, at some point during their larval or adult stages, five distinctive physical characteristics (synapomorphies) that distinguish them from other taxa. These five synapomorphies are a notochord, a hollow dorsal nerve cord, an endostyle or thyroid, pharyngeal slits, and a post-anal tail. The name "chordate" comes from the first of these synapomorphies, the notochord, which plays a significant role in chordate body plan structuring and movements. Chordates are also bilaterally symmetric, have a coelom, possess a closed circulatory system, and exhibit metameric segmentation.

Vertebrates are deuterostomal animals with bony or cartilaginous axial endoskeleton — known as the vertebral column, spine or backbone — around and along the spinal cord, including all fish, amphibians, reptiles, birds and mammals. The vertebrates consist of all the taxa within the subphylum Vertebrata and represent the overwhelming majority of the phylum Chordata, with currently about 69,963 species described.

Hagfish, of the class Myxini and order Myxiniformes, are eel-shaped jawless fish. Hagfish are the only known living animals that have a skull but no vertebral column, although they do have rudimentary vertebrae. Hagfish are marine predators and scavengers who can defend themselves against other larger predators by releasing copious amounts of slime from mucous glands in their skin.

Agnatha is an infraphylum of jawless fish in the phylum Chordata, subphylum Vertebrata, consisting of both living (cyclostomes) and extinct species. Among recent animals, cyclostomes are sister to all vertebrates with jaws, known as gnathostomes.

Gnathostomata are the jawed vertebrates. Gnathostome diversity comprises roughly 60,000 species, which accounts for 99% of all living vertebrates, including humans. In addition to opposing jaws, living gnathostomes have true teeth, paired appendages, the elastomeric protein of elastin, and a horizontal semicircular canal of the inner ear, along with physiological and cellular anatomical characters such as the myelin sheaths of neurons, and an adaptive immune system that has the discrete lymphoid organs of spleen and thymus, and uses V(D)J recombination to create antigen recognition sites, rather than using genetic recombination in the variable lymphocyte receptor gene.

Fish anatomy is the study of the form or morphology of fish. It can be contrasted with fish physiology, which is the study of how the component parts of fish function together in the living fish. In practice, fish anatomy and fish physiology complement each other, the former dealing with the structure of a fish, its organs or component parts and how they are put together, such as might be observed on the dissecting table or under the microscope, and the latter dealing with how those components function together in living fish.

A craniate is a member of the Craniata, a proposed clade of chordate animals with a skull of hard bone or cartilage. Living representatives are the Myxini (hagfishes), Hyperoartia, and the much more numerous Gnathostomata. Formerly distinct from vertebrates by excluding hagfish, molecular and anatomical research in the 21st century has led to the reinclusion of hagfish as vertebrates, making living craniates synonymous with living vertebrates.
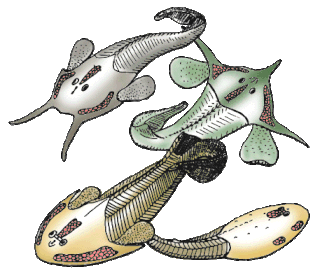
Ostracoderms are the armored jawless fish of the Paleozoic Era. The term does not often appear in classifications today because it is paraphyletic and thus does not correspond to one evolutionary lineage. However, the term is still used as an informal way of loosely grouping together the armored jawless fishes.

The class Osteostraci is an extinct taxon of bony-armored jawless fish, termed "ostracoderms", that lived in what is now North America, Europe and Russia from the Middle Silurian to Late Devonian.

Pteraspidomorphi is an extinct class of early jawless fish. They have long been regarded as closely related or even ancestral to jawed vertebrates, but the few characteristics they share with the latter are now considered as basal traits for all vertebrates.
Cyclostome is a biological term used in a few different senses:

Marine vertebrates are vertebrates that live in marine environments. These are the marine fish and the marine tetrapods. Vertebrates are a subphylum of chordates that have a vertebral column (backbone). The vertebral column provides the central support structure for an internal skeleton. The internal skeleton gives shape, support, and protection to the body and can provide a means of anchoring fins or limbs to the body. The vertebral column also serves to house and protect the spinal cord that lies within the column.

Metaspriggina is a genus of chordate initially known from two specimens in the Middle Cambrian Burgess Shale and 44 specimens found in 2012 at the Marble Canyon bed in Kootenay National Park.

Lampreys are a group of jawless fish comprising the order Petromyzontiformes. The adult lamprey is characterized by a toothed, funnel-like sucking mouth. The common name "lamprey" is probably derived from Latin lampetra, which may mean "stone licker", though the etymology is uncertain. Lamprey is sometimes seen for the plural form.

Most bony fishes have two sets of jaws made mainly of bone. The primary oral jaws open and close the mouth, and a second set of pharyngeal jaws are positioned at the back of the throat. The oral jaws are used to capture and manipulate prey by biting and crushing. The pharyngeal jaws, so-called because they are positioned within the pharynx, are used to further process the food and move it from the mouth to the stomach.

The evolution of fish began about 530 million years ago during the Cambrian explosion. It was during this time that the early chordates developed the skull and the vertebral column, leading to the first craniates and vertebrates. The first fish lineages belong to the Agnatha, or jawless fish. Early examples include Haikouichthys. During the late Cambrian, eel-like jawless fish called the conodonts, and small mostly armoured fish known as ostracoderms, first appeared. Most jawless fish are now extinct; but the extant lampreys may approximate ancient pre-jawed fish. Lampreys belong to the Cyclostomata, which includes the extant hagfish, and this group may have split early on from other agnathans.

Priscomyzon riniensis is an extinct lamprey that lived some 360 million years ago during the Famennian in a marine or estuarine environment in South Africa. This small agnathan is anatomically similar to the Mazon Creek lampreys, but is some 35 million years older. Its key developments included the first known large oral disc, circumoral teeth and a branchial basket.

Romundina is a small, heavily armored extinct genus of acanthothoracid placoderms which lived in shallow marine environments in the early Devonian (Lochkovian). The name Romundina honors Canadian geologist and paleontologist Dr. Rómundur (Raymond) Thorsteinsson of Calgary, Alberta, Canada. Romundina are believed to have lived on Earth between 400 and 419 million years ago. The closest known relative to Romundina is the acanthothoracid Radotina. The type and only described species is R. stellina.

Myxinikela is an extinct genus of stem-hagfish known from the Late Carboniferous of Illinois, USA. It is the earliest definitive hagfish known from fossil remains, and one of only two alongside the Cretaceous crown-group hagfish Tethymyxine.
Fishes are a paraphyletic group and for this reason, the class Pisces seen in older reference works is no longer used in formal taxonomy. Traditional classification divides fish into three extant classes, and with extinct forms sometimes classified within those groups, sometimes as their own classes:

