
Glutamic acid is an α-amino acid that is used by almost all living beings in the biosynthesis of proteins. It is a non-essential nutrient for humans, meaning that the human body can synthesize enough for its use. It is also the most abundant excitatory neurotransmitter in the vertebrate nervous system. It serves as the precursor for the synthesis of the inhibitory gamma-aminobutyric acid (GABA) in GABAergic neurons.

The GABA receptors are a class of receptors that respond to the neurotransmitter gamma-aminobutyric acid (GABA), the chief inhibitory compound in the mature vertebrate central nervous system. There are two classes of GABA receptors: GABAA and GABAB. GABAA receptors are ligand-gated ion channels ; whereas GABAB receptors are G protein-coupled receptors, also called metabotropic receptors.

Aromatic L-amino acid decarboxylase, also known as DOPA decarboxylase (DDC), tryptophan decarboxylase, and 5-hydroxytryptophan decarboxylase, is a lyase enzyme, located in region 7p12.2-p12.1.

Pyridoxal phosphate (PLP, pyridoxal 5'-phosphate, P5P), the active form of vitamin B6, is a coenzyme in a variety of enzymatic reactions. The International Union of Biochemistry and Molecular Biology has catalogued more than 140 PLP-dependent activities, corresponding to ~4% of all classified activities. The versatility of PLP arises from its ability to covalently bind the substrate, and then to act as an electrophilic catalyst, thereby stabilizing different types of carbanionic reaction intermediates.

Calcineurin (CaN) is a calcium and calmodulin dependent serine/threonine protein phosphatase. It activates the T cells of the immune system and can be blocked by drugs. Calcineurin activates nuclear factor of activated T cell cytoplasmic (NFATc), a transcription factor, by dephosphorylating it. The activated NFATc is then translocated into the nucleus, where it upregulates the expression of interleukin 2 (IL-2), which, in turn, stimulates the growth and differentiation of the T cell response. Calcineurin is the target of a class of drugs called calcineurin inhibitors, which include ciclosporin, voclosporin, pimecrolimus and tacrolimus.

Aspartate transaminase (AST) or aspartate aminotransferase, also known as AspAT/ASAT/AAT or (serum) glutamic oxaloacetic transaminase, is a pyridoxal phosphate (PLP)-dependent transaminase enzyme that was first described by Arthur Karmen and colleagues in 1954. AST catalyzes the reversible transfer of an α-amino group between aspartate and glutamate and, as such, is an important enzyme in amino acid metabolism. AST is found in the liver, heart, skeletal muscle, kidneys, brain, red blood cells and gall bladder. Serum AST level, serum ALT level, and their ratio are commonly measured clinically as biomarkers for liver health. The tests are part of blood panels.

Glutamate dehydrogenase is an enzyme observed in both prokaryotes and eukaryotic mitochondria. The aforementioned reaction also yields ammonia, which in eukaryotes is canonically processed as a substrate in the urea cycle. Typically, the α-ketoglutarate to glutamate reaction does not occur in mammals, as glutamate dehydrogenase equilibrium favours the production of ammonia and α-ketoglutarate. Glutamate dehydrogenase also has a very low affinity for ammonia, and therefore toxic levels of ammonia would have to be present in the body for the reverse reaction to proceed. However, in brain, the NAD+/NADH ratio in brain mitochondria encourages oxidative deamination. In bacteria, the ammonia is assimilated to amino acids via glutamate and aminotransferases. In plants, the enzyme can work in either direction depending on environment and stress. Transgenic plants expressing microbial GLDHs are improved in tolerance to herbicide, water deficit, and pathogen infections. They are more nutritionally valuable.
Slowly evolving immune-mediated diabetes, or latent autoimmune diabetes in adults (LADA), is a form of diabetes that exhibits clinical features similar to both type 1 diabetes (T1D) and type 2 diabetes (T2D), and is sometimes referred to as type 1.5 diabetes. It is an autoimmune form of diabetes, similar to T1D, but patients with LADA often show insulin resistance, similar to T2D, and share some risk factors for the disease with T2D. Studies have shown that LADA patients have certain types of antibodies against the insulin-producing cells, and that these cells stop producing insulin more slowly than in T1D patients. Since many people develop the disease later in life, it is often misdiagnosed as type 2 diabetes.

N-Acetylaspartylglutamic acid is a peptide neurotransmitter and the third-most-prevalent neurotransmitter in the mammalian nervous system. NAAG consists of N-acetylaspartic acid (NAA) and glutamic acid coupled via a peptide bond.
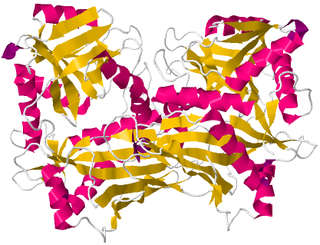
Branched-chain amino acid aminotransferase (BCAT), also known as branched-chain amino acid transaminase, is an aminotransferase enzyme (EC 2.6.1.42) which acts upon branched-chain amino acids (BCAAs). It is encoded by the BCAT2 gene in humans. The BCAT enzyme catalyzes the conversion of BCAAs and α-ketoglutarate into branched chain α-keto acids and glutamate.

The enzyme Acid-Induced Arginine Decarboxylase (AdiA), also commonly referred to as arginine decarboxylase, catalyzes the conversion of L-arginine into agmatine and carbon dioxide. The process consumes a proton in the decarboxylation and employs a pyridoxal-5'-phosphate (PLP) cofactor, similar to other enzymes involved in amino acid metabolism, such as ornithine decarboxylase and glutamine decarboxylase. It is found in bacteria and virus, though most research has so far focused on forms of the enzyme in bacteria. During the AdiA catalyzed decarboxylation of arginine, the necessary proton is consumed from the cell cytoplasm which helps to prevent the over-accumulation of protons inside the cell and serves to increase the intracellular pH. Arginine decarboxylase is part of an enzymatic system in Escherichia coli, Salmonella Typhimurium, and methane-producing bacteria Methanococcus jannaschii that makes these organisms acid resistant and allows them to survive under highly acidic medium.

In enzymology, 4-aminobutyrate transaminase, also called GABA transaminase or 4-aminobutyrate aminotransferase, or GABA-T, is an enzyme that catalyzes the chemical reaction:

The GABAA beta-2 subunit is a protein that in humans is encoded by the GABRB2 gene. It combines with other subunits to form the ionotropic GABAA receptors. GABA system is the major inhibitory system in the brain, and its dominant GABAA receptor subtype is composed of α1, β2, and γ2 subunits with the stoichiometry of 2:2:1, which accounts for 43% of all GABAA receptors. Alternative splicing of the GABRB2 gene leads at least to four isoforms, viz. β2-long (β2L) and β2-short. Alternatively spliced variants displayed similar but non-identical electrophysiological properties. GABRB2 is subjected to positive selection and known to be both an alternative splicing and a recombination hotspot; it is regulated via epigenetic regulation including imprinting and gene and promoter methylation GABRB2 has been associated with a number of neuropsychiatric disorders, and found to display altered expression in cancer.

Receptor-type tyrosine-protein phosphatase-like N, also called "IA-2", is an enzyme that in humans is encoded by the PTPRN gene.

GABA transporter 1 (GAT1) also known as sodium- and chloride-dependent GABA transporter 1 is a protein that in humans is encoded by the SLC6A1 gene and belongs to the solute carrier 6 (SLC6) family of transporters. It mediates gamma-aminobutyric acid's translocation from the extracellular to intracellular spaces within brain tissue and the central nervous system as a whole.
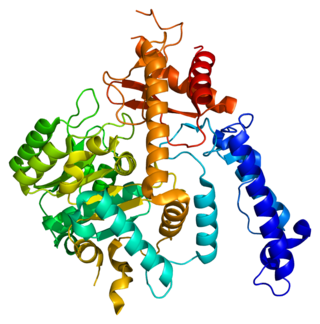
Glutamate decarboxylase 2 is an enzyme that in humans is encoded by the GAD2 gene.

Glutamate decarboxylase 1 (GAD67), also known as GAD1, is a human gene.
In biochemistry, the glutamate–glutamine cycle is a cyclic metabolic pathway which maintains an adequate supply of the neurotransmitter glutamate in the central nervous system. Neurons are unable to synthesize either the excitatory neurotransmitter glutamate, or the inhibitory GABA from glucose. Discoveries of glutamate and glutamine pools within intercellular compartments led to suggestions of the glutamate–glutamine cycle working between neurons and astrocytes. The glutamate/GABA–glutamine cycle is a metabolic pathway that describes the release of either glutamate or GABA from neurons which is then taken up into astrocytes. In return, astrocytes release glutamine to be taken up into neurons for use as a precursor to the synthesis of either glutamate or GABA.

Autoimmune encephalitis (AIE) is a type of encephalitis, and one of the most common causes of noninfectious encephalitis. It can be triggered by tumors, infections, or it may be cryptogenic. The neurological manifestations can be either acute or subacute and usually develop within six weeks. The clinical manifestations include behavioral and psychiatric symptoms, autonomic disturbances, movement disorders, and seizures.
Fernando Garcia de Mello is a renowned neurochemist from Brazil. He obtained his degree in Biochemistry in 1968 from the State University of Rio de Janeiro. Fernando Mello started his scientific training as an undergraduate student at the Brazilian National Institute of Cancer, and later at the Institute of Biophysics from the Federal University of Rio de Janeiro, being mentored by dr. Firmino de Castro, which greatly influenced him to have a more humanistic approach towards the students that he would train. It was only during his post-doc period (1973-1976) at the National Institutes of Health under supervision of dr. Marshall Warren Nirenberg that Mello began his research in Neurochemistry, using the embryonary Retina as a model for his investigations.




