Articles related to anatomy include:

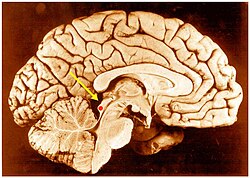
The brainstem is the stalk-like part of the brain that interconnects the cerebrum and diencephalon with the spinal cord. In the human brain, the brainstem is composed of the midbrain, the pons, and the medulla oblongata. The midbrain is continuous with the thalamus of the diencephalon through the tentorial notch.

In neuroanatomy, the lateral geniculate nucleus is a structure in the thalamus and a key component of the mammalian visual pathway. It is a small, ovoid, ventral projection of the thalamus where the thalamus connects with the optic nerve. There are two LGNs, one on the left and another on the right side of the thalamus. In humans, both LGNs have six layers of neurons alternating with optic fibers.
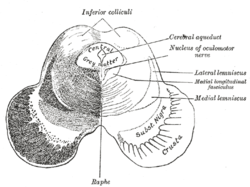
The midbrain or mesencephalon is the rostral-most portion of the brainstem connecting the diencephalon and cerebrum with the pons. It consists of the cerebral peduncles, tegmentum, and tectum.

The spinothalamic tract is a part of the anterolateral system or the ventrolateral system, a sensory pathway to the thalamus. From the ventral posterolateral nucleus in the thalamus, sensory information is relayed upward to the somatosensory cortex of the postcentral gyrus.

The auditory system is the sensory system for the sense of hearing. It includes both the sensory organs and the auditory parts of the sensory system.

In neuroanatomy, the medial lemniscus, also known as Reil's band or Reil's ribbon, is a large ascending bundle of heavily myelinated axons that decussate (cross) in the brainstem, specifically in the medulla oblongata. The medial lemniscus is formed by the crossings of the internal arcuate fibers. The internal arcuate fibers are composed of axons of nucleus gracilis and nucleus cuneatus. The axons of the nucleus gracilis and nucleus cuneatus in the medial lemniscus have cell bodies that lie contralaterally.

The lateral lemniscus is a tract of axons in the brainstem that carries information about sound from the cochlear nucleus to various brainstem nuclei and ultimately the contralateral inferior colliculus of the midbrain. Three distinct, primarily inhibitory, cellular groups are located interspersed within these fibers, and are thus named the nuclei of the lateral lemniscus.

In neuroanatomy, the superior colliculus is a structure lying on the roof of the mammalian midbrain. In non-mammalian vertebrates, the homologous structure is known as the optic tectum or optic lobe. The adjective form tectal is commonly used for both structures.

In neuroanatomy, the pretectal area, or pretectum, is a midbrain structure composed of seven nuclei and comprises part of the subcortical visual system. Through reciprocal bilateral projections from the retina, it is involved primarily in mediating behavioral responses to acute changes in ambient light such as the pupillary light reflex, the optokinetic reflex, and temporary changes to the circadian rhythm. In addition to the pretectum's role in the visual system, the anterior pretectal nucleus has been found to mediate somatosensory and nociceptive information.
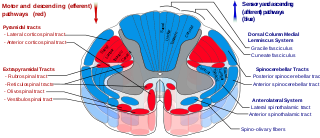
The spinocerebellar tract is a nerve tract originating in the spinal cord and terminating in the same side (ipsilateral) of the cerebellum.

The medial geniculate nucleus (MGN) or medial geniculate body (MGB) is part of the auditory thalamus and represents the thalamic relay between the inferior colliculus (IC) and the auditory cortex (AC). It is made up of a number of sub-nuclei that are distinguished by their neuronal morphology and density, by their afferent and efferent connections, and by the coding properties of their neurons. It is thought that the MGN influences the direction and maintenance of attention.

In the brain, the corpora quadrigemina are the four colliculi—two inferior, two superior—located on the tectum of the dorsal aspect of the midbrain. They are respectively named the inferior and superior colliculus.

The cochlear nuclear (CN) complex comprises two cranial nerve nuclei in the human brainstem, the ventral cochlear nucleus (VCN) and the dorsal cochlear nucleus (DCN). The ventral cochlear nucleus is unlayered whereas the dorsal cochlear nucleus is layered. Auditory nerve fibers, fibers that travel through the auditory nerve carry information from the inner ear, the cochlea, on the same side of the head, to the nerve root in the ventral cochlear nucleus. At the nerve root the fibers branch to innervate the ventral cochlear nucleus and the deep layer of the dorsal cochlear nucleus. All acoustic information thus enters the brain through the cochlear nuclei, where the processing of acoustic information begins. The outputs from the cochlear nuclei are received in higher regions of the auditory brainstem.

The superior olivary complex (SOC) or superior olive is a collection of brainstem nuclei that is located in pons, functions in multiple aspects of hearing and is an important component of the ascending and descending auditory pathways of the auditory system. The SOC is intimately related to the trapezoid body: most of the cell groups of the SOC are dorsal to this axon bundle while a number of cell groups are embedded in the trapezoid body. Overall, the SOC displays a significant interspecies variation, being largest in bats and rodents and smaller in primates.

The interaural time difference when concerning humans or animals, is the difference in arrival time of a sound between two ears. It is important in the localization of sounds, as it provides a cue to the direction or angle of the sound source from the head. If a signal arrives at the head from one side, the signal has further to travel to reach the far ear than the near ear. This pathlength difference results in a time difference between the sound's arrivals at the ears, which is detected and aids the process of identifying the direction of sound source.

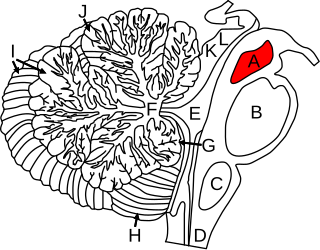
Cerebellar peduncles connect the cerebellum to the brain stem. There are six cerebellar peduncles in total, three on each side:

In the human brain, the superior cerebellar peduncle is a paired structure of white matter that connects the cerebellum to the midbrain. It consists mainly of efferent fibers, the cerebellothalamic tract that runs from a cerebellar hemisphere to the contralateral thalamus, and the cerebellorubral tract that runs from a cerebellar hemisphere to the red nucleus. It also contains afferent tracts, most prominent of which is the ventral spinocerebellar tract. Other afferent tracts are the trigeminothalamic fibers, tectocerebellar fibers, and noradrenergic fibers from the locus coeruleus. The superior peduncle emerges from the upper and medial parts of the white matter of each hemisphere and is placed under cover of the upper part of the cerebellum.

In the ventral cochlear nucleus (VCN), auditory nerve fibers enter the brain via the nerve root in the VCN. The ventral cochlear nucleus is divided into the anterior ventral (anteroventral) cochlear nucleus (AVCN) and the posterior ventral (posteroventral) cochlear nucleus (PVCN). In the VCN, auditory nerve fibers bifurcate, the ascending branch innervates the AVCN and the descending branch innervates the PVCN and then continue to the dorsal cochlear nucleus. The orderly innervation by auditory nerve fibers gives the AVCN a tonotopic organization along the dorsoventral axis. Fibers that carry information from the apex of the cochlea that are tuned to low frequencies contact neurons in the ventral part of the AVCN; those that carry information from the base of the cochlea that are tuned to high frequencies contact neurons in the dorsal part of the AVCN. Several populations of neurons populate the AVCN. Bushy cells receive input from auditory nerve fibers through particularly large endings called end bulbs of Held. They contact stellate cells through more conventional boutons.