Interleukin 17 family (IL17 family) is a family of pro-inflammatory cystine knotcytokines.[2] They are produced by a group of T helper cell known as T helper 17 cell in response to their stimulation with IL-23. Originally, Th17 was identified in 1993 by Rouvier et al. who isolated IL17A transcript from a rodentT-cellhybridoma.[3] The protein encoded by IL17A is a founding member of IL-17 family (see below). IL17A protein exhibits a high homology with a viral IL-17-like protein (O40633) encoded in the genome of T-lymphotropic rhadinovirusHerpesvirus saimiri. In rodents, IL-17A is often referred to as CTLA8.[4]
The biologically active IL-17 interacts with type I cell surface receptor IL-17R. In turn, there are at least three variants of IL-17R referred to as IL17RA, IL17RB, and IL17RC.[5] After binding to the receptor, IL-17 activates several signalling cascades that, in turn, lead to the induction of chemokines. Acting as chemoattractants, these chemokines recruit the immune cells, such as monocytes and neutrophils to the site of inflammation. Typically, the signaling events mentioned above follow an invasion of the body by pathogens. Promoting the inflammation, IL-17 acts in concert with tumor necrosis factor and interleukin-1.[6][7] Moreover, an activation of IL-17 signalling is often observed in the pathogenesis of various autoimmune disorders, such as psoriasis.[8]
Family members
The IL-17 family in humans comprises IL17A (sometimes confusingly called "IL-17"), IL17B, IL17C, IL17D, IL17E and IL17F. IL-17E is also known as IL-25. All members of the IL-17 family have a similar protein structure. Their protein sequences contain four highly conserved cysteine residues. These conserved cysteine residues are critical to the right 3-dimensional shape of the entire protein molecule. To the reference, the members of the IL-17 family do not exhibit a significant sequence homology with other cytokines. Among IL-17 family members, the IL-17F isoforms 1 and 2 (ML-1) have the highest sequence homology with IL-17A (55 and 40%, respectively). They follow by IL-17B, which has 29% similarity to IL-17A, IL-17D (25%), IL-17C (23%), and IL-17E (17%). In mammals, the sequences of these cytokines are highly conserved. For instance, the sequence homology between the corresponding human and mouse proteins is usually between 62–88%.[9]
Function
Numerous immune regulatory functions have been reported for the IL-17 family of cytokines, presumably due to their induction of many immune signaling molecules. The most notable role of IL-17 is its involvement in inducing and mediating proinflammatory responses. IL-17 is commonly associated with allergic responses. IL-17 induces the production of many other cytokines (such as IL-6, G-CSF, GM-CSF, IL-1β, TGF-β, TNF-α), chemokines (including IL-8, GRO-α, and MCP-1), and prostaglandins (e.g., PGE2) from many cell types (fibroblasts, endothelial cells, epithelial cells, keratinocytes, and macrophages). IL-17 acts with IL-22 (produced mainly by T helper 22 cells in humans, but by T helper 17 cell in mice) to induce expression of antimicrobial peptide by keratinocytes.
The release of cytokines causes many functions, such as airway remodeling, a characteristic of IL-17 responses. The increased expression of chemokines attracts other cells including neutrophils but not eosinophils. IL-17 function is also essential to a subset of CD4+ T-Cells called T helper 17 (Th17) cells. As a result of these roles, the IL-17 family has been linked to many immune-related/autoimmune diseases including rheumatoid arthritis, asthma, lupus, allograft rejection, anti-tumour immunity and recently psoriasis,[10]multiple sclerosis,[11] and intracerebral hemorrhage.[12]
Gene expression
The gene for human IL-17A is 1874 base pairs long[13] and was cloned from CD4+ T cells. Each member of the IL-17 family has a distinct pattern of cellular expression. The expression of IL-17A and IL-17F appear to be restricted to a small group of activated T cells, and upregulated during inflammation. IL-17B is expressed in several peripheral tissues and immune tissues. IL-17C is also highly upregulated in inflammatory conditions, although in resting conditions is low in abundance. IL-17D is highly expressed in the nervous system and in skeletal muscle and IL-17E is found at low levels in various peripheral tissues.[10]
Much progress has been made in the understanding of the regulation of IL-17. At first, Aggarwal et al. showed that production of IL-17 was dependent on IL-23.[14] Later, a Korean group discovered that STAT3 and NF-κB signalling pathways are required for this IL-23-mediated IL-17 production.[15] Consistent with this finding, Chen et al. showed that another molecule, SOCS3, plays an important role in IL-17 production.[16] In the absence of SOCS3, IL-23-induced STAT3 phosphorylation is enhanced, and phosphorylated STAT3 binds to the promoter regions of both IL-17A and IL-17F increasing their gene activity. In contrast, some scientists believe IL-17 induction is independent of IL-23. Several groups have identified ways to induce IL-17 production both in vitro[17] and in vivo[18][19] by distinct cytokines, called TGF-β and IL-6, without the need for IL-23.[17][18][19] Although IL-23 is not required for IL-17 expression in this situation, IL-23 may play a role in promoting survival and/or proliferation of the IL-17 producing T-cells. Recently, Ivanov et al. found that the thymus specific nuclear receptor, ROR-γ, directs differentiation of IL-17-producing T cells.[20]
Structure
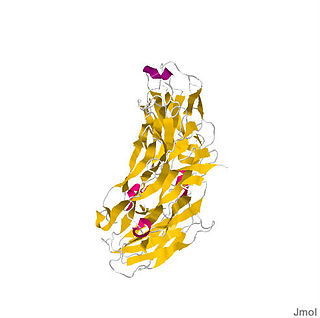
IL-17(A) is a 155-amino acid protein that is a disulfide-linked, homodimeric, secreted glycoprotein with a molecular mass of 35 kDa.[9] Each subunit of the homodimer is approximately 15-20 KDa. The structure of IL-17 consists of a signal peptide of 23 amino acids (aa) followed by a 123-aa chain region characteristic of the IL-17 family. An N-linked glycosylation site on the protein was first identified after purification of the protein revealed two bands, one at 15 KDa and another at 20 KDa. Comparison of different members of the IL-17 family revealed four conserved cysteines that form two disulfide bonds.[13] IL-17 is unique in that it bears no resemblance to other known interleukins. Furthermore, IL-17 bears no resemblance to any other known proteins or structural domains.[10]
The crystal structure of IL-17F, which is 50% homologous to IL-17A, revealed that IL-17F is structurally similar to the cystine knot family of proteins that includes the neurotrophins. The cystine knot fold is characterized by two sets of paired β-strands stabilized by three disulfide interactions. However, in contrast to the other cystine knot proteins, IL-17F lacks the third disulfide bond. Instead, a serine replaces the cysteine at this position. This unique feature is conserved in the other IL-17 family members. IL-17F also dimerizes in a fashion similar to nerve growth factor (NGF) and other neurotrophins.[1]
Role in psoriasis
Recent work suggests the IL-23/IL-17 pathway plays a major role in the autoimmune disorder psoriasis.[8][21][22] In this condition, immune cells react to inflammatory molecules released within the skin around the joints and scalp.[21] This response causes the epidermal cells to recycle more rapidly than usual, which leads to the formation of red, scaly lesions and chronic skin inflammation.[22][23] Analysis of biopsies taken from lesions of psoriasis patients show an enrichment of cytotoxic T cells and neutrophils containing IL-17.[21][24][25] This indicates an excessive infiltration of pro-inflammatory immune cells and IL-17 cytokines are associated with the development of psoriasis.
Studies conducted in mice demonstrate that removing either IL-23 or IL-17 decreases the progression of psoriasis.[26][27] Mice injected with monoclonal antibodies targeting IL-17 blocked, or neutralized, down-stream signaling of this cytokine and decreased epidermal hyperplasia.[26] Similarly, mice genetically modifying to not express IL-23 or IL-17 receptors significantly reduced psoriatic lesion development upon stimulation with the lesion-causing tumor promoter 12-O-tetradecanoylphorbol-13-acetate.[8]
IL-17 promotes psoriasis by contributing to the inflammatory response that damages and overturns the keratinocyte cells of the epidermal layer.[21][27] Inflammation begins with keratinocyte cells entering the final stages of their cell cycle, which activates immature dendritic cells (DC).[28] Cytokines released from DCs stimulate dying keratinocytes to secrete TNF-alpha, IL-1 and IL-6 leading to the chemotaxis of T cells, natural killer cells and monocytes to the epidermis.[23] These cells release IL-23 which induce Th17 cells to produce IL-17.[24]
IL-17 interaction with IL-17RA receptors, abundant on the keratinocyte cell surface, incite epidermal cells to increase expression of IL-6, antimicrobial peptides, IL-8 and CCL20.[8][22][27] Increased concentration of IL-6 alters the epidermal environment by decreasing the ability of T regulatory cells to control the behavior of Th17 cells.[24] Reduced regulation allows uninhibited proliferation of Th17 cells and production of IL-17 in psoriatic lesions, augmenting IL-17 signaling.[24] Antimicrobial peptides and IL-8 attract neutrophils to the site of injury where these cells remove damaged and inflamed keratinocyte cells.[21][25][27] New immature DCs are also recruited by CCL20 via chemotaxis where their activation restarts and amplifies the cycle of inflammation.[24][25] IL-17 and additional cytokines released from the influx of neutrophils, T and dendritic cells mediate effects on localized leukocytes and keratinocytes that supports the progression of psoriasis by inciting chronic inflammation.[24]
Role in asthma
The IL-17F gene was discovered in 2001 and is located on chromosome 6p12. Notably, among this family, IL-17F has been well characterized both in vitro and in vivo and has been shown to have a pro-inflammatory role in asthma. IL-17F is clearly expressed in the airway of asthmatics and its expression level is correlated with disease severity. Moreover, a coding region variant (H161R) of the IL-17F gene is inversely associated with asthma and encodes an antagonist for the wild-type IL-17F. IL-17F is able to induce several cytokines, chemokines and adhesion molecules in bronchial epithelial cells, vein endothelial cells, fibroblasts and eosinophils. IL-17F utilizes IL-17RA and IL-17RC as its receptors and activates the MAP kinase-related pathway. IL-17F is derived from several cell types such as Th17 cells, mast cells and basophils, and shows a wide tissue expression pattern including lung. Overexpression of IL-17F gene in the airway of mice is associated with airway neutrophilia, the induction of many cytokines, an increase in airway hyperreactivity, and mucus hypersecretion. Hence, IL-17F may have a crucial role in allergic airway inflammation and have important therapeutic implications in asthma.[29]
Based on emerging evidence from animal models, IL-17 has been suggested as a target for anti-inflammatory therapies to improve recovery post-stroke[36] and to reduce the formation of skin cancer.[37] IL-17 has also been implicated in multiple sclerosis.[11]
The active form of vitamin D has been found to 'severely impair' [38] production of the IL-17 and IL-17F cytokines by Th17 cells.
The IL-17 receptor family consists of five, broadly distributed receptors (IL-17RA, B, C, D and E) that present with individual ligand specificities. Within this family of receptors, IL-17RA is the best-described. IL-17RA binds both IL-17A and IL-17F and is expressed in multiple tissues: vascular endothelial cells, peripheral T cells, B cell lineages, fibroblast, lung, myelomonocytic cells, and marrow stromal cells.[9][39][2] Signal transduction for both IL-17A and IL-17F requires the presence of a heterodimeric complex consisting of both IL-17RA and IL-17RC and the absence of either receptor results in ineffective signal transduction. This pattern is reciprocated for other members of the IL-17 family such as IL-17E, which requires an IL-17RA-IL-17RB complex (also known as IL-17Rh1, IL-17BR or IL-25R) for effective function.[40]
Another member of this receptor family, IL-17RB, binds both IL-17B and IL-17E.[9][2] Furthermore, it is expressed in the kidney, pancreas, liver, brain, and intestine.[9] IL-17RC is expressed by the prostate, cartilage, kidney, liver, heart, and muscle, and its gene may undergo alternate splicing to produce a soluble receptor in addition to its cell membrane-bound form. In a similar manner, the gene for IL-17RD may undergo alternative splicing to yield a soluble receptor. This feature may allow these receptors to inhibit the stimulatory effects of their yet-undefined ligands.[9][2] The least-described of these receptors, IL-17RE, is known to be expressed in the pancreas, brain, and prostate.[9]
Signal transduction by these receptors is as diverse as their distribution. These receptors do not exhibit a significant similarity in extracellular or intracellular amino acid sequence when compared to other cytokine receptors.[39] Transcription factors such as TRAF6, JNK, Erk1/2, p38, AP-1 and NF-κB have been implicated in IL-17 mediated signaling in a stimulation-dependent, tissue-specific manner.[39][2][41] Other signaling mechanisms have also been proposed, but more work is needed to fully elucidate the true signaling pathways used by these diverse receptors.
Related Research Articles
Interleukin 12 (IL-12) is an interleukin that is naturally produced by dendritic cells, macrophages, neutrophils, helper T cells and human B-lymphoblastoid cells (NC-37) in response to antigenic stimulation. IL-12 belongs to the family of interleukin-12. IL-12 family is unique in comprising the only heterodimeric cytokines, which includes IL-12, IL-23, IL-27 and IL-35. Despite sharing many structural features and molecular partners, they mediate surprisingly diverse functional effects.
Interleukin-23 subunit alpha is a protein that in humans is encoded by the IL23A gene. The protein is also known as IL-23p19. It is one of the two subunits of the cytokine Interleukin-23.
Interleukin-26 (IL-26) is a protein that in humans is encoded by the IL26 gene.
Interleukin-25 (IL-25) – also known as interleukin-17E (IL-17E) – is a protein that in humans is encoded by the IL25 gene on chromosome 14. IL-25 was discovered in 2001 and is made up of 177 amino acids.
Interleukin-22 (IL-22) is a protein that in humans is encoded by the IL22 gene.
Interleukin 20 (IL20) is a protein that is in humans encoded by the IL20 gene which is located in close proximity to the IL-10 gene on the 1q32 chromosome. IL-20 is a part of an IL-20 subfamily which is a part of a larger IL-10 family.
Interleukin 19 (IL-19) is an immunosuppressive protein that belongs to the IL-10 cytokine subfamily.
T helper 17 cells (Th17) are a subset of pro-inflammatory T helper cells defined by their production of interleukin 17 (IL-17). They are related to T regulatory cells and the signals that cause Th17s to actually inhibit Treg differentiation. However, Th17s are developmentally distinct from Th1 and Th2 lineages. Th17 cells play an important role in maintaining mucosal barriers and contributing to pathogen clearance at mucosal surfaces; such protective and non-pathogenic Th17 cells have been termed as Treg17 cells.
Interleukin 17 receptor A, also known as IL17RA and CDw217, is a human gene.
Interleukin 20 receptors (IL20R) belong to the IL-10 family. IL20R are involved in both pro-inflammatory and anti-inflammatory immune response. There are two types of IL20R: Type I and Type II.
Interleukin-17A is a protein that in humans is encoded by the IL17A gene. In rodents, IL-17A used to be referred to as CTLA8, after the similarity with a viral gene.
Interleukin-17 receptor (IL-17R) is a cytokine receptor which belongs to new subfamily of receptors binding proinflammatory cytokine interleukin 17A, a member of IL-17 family ligands produced by T helper 17 cells (Th17). IL-17R family consists of 5 members: IL-17RA, IL-17RB, IL-17RC, IL-17RD and IL-17RE. Functional IL-17R is a transmembrane receptor complex usually consisting of one IL-17RA, which is a founding member of the family, and second other family subunit, thus forming heteromeric receptor binding different ligands. IL-17A, a founding member of IL-17 ligand family binds to heteromeric IL-17RA/RC receptor complex. IL-17RB binds preferentially IL-17B and IL-17E and heteromeric IL-17RA/RE complex binds IL-17C. However, there is still unknown ligand for IL-17RD. The first identified member IL-17RA is located on human chromosome 22, whereas other subunits IL-17RB to IL-17RD are encoded within human chromosome 3.
The Interleukin-1 family is a group of 11 cytokines that plays a central role in the regulation of immune and inflammatory responses to infections or sterile insults.
Interleukin 23 (IL-23) is a heterodimeric cytokine composed of an IL-12B (IL-12p40) subunit and an IL-23A (IL-23p19) subunit. IL-23 is part of the IL-12 family of cytokines. The functional receptor for IL-23 consists of a heterodimer between IL-12Rβ1 and IL-23R.
Interleukin 36, or IL-36, is a group of cytokines in the IL-1 family with pro-inflammatory effects. The role of IL-36 in inflammatory diseases is under investigation.
Interleukin 17 receptor D is a protein that in humans is encoded by the IL17RD gene.
Type 3 innate lymphoid cells (ILC3) are immune cells from the lymphoid lineage that are part of the innate immune system. These cells participate in innate mechanisms on mucous membranes, contributing to tissue homeostasis, host-commensal mutualism and pathogen clearance. They are part of a heterogeneous group of innate lymphoid cells, which is traditionally divided into three subsets based on their expression of master transcription factors as well as secreted effector cytokines - ILC1, ILC2 and ILC3.
Interleukin 17F (IL-17F) is signaling protein that is in human is encoded by the IL17F gene and is considered a pro-inflammatory cytokine. This protein belongs to the interleukin 17 family and is mainly produced by the T helper 17 cells after their stimulation with interleukin 23. However, IL-17F can be also produced by a wide range of cell types, including innate immune cells and epithelial cells.
Th22 cells are subpopulation of CD4+ T cells that produce interleukin-22 (IL-22). They play a role in the protective mechanisms against variety of bacterial pathogens, tissue repair and wound healing, and also in pathologic processes, including inflammations, autoimmunity, tumors, and digestive organs damages.
Act 1 adaptor protein is an essential intermediate in the interleukin-17 pathway. The IL-17 protein is a pro-inflammatory cytokine important for tissue inflammation in host defense against infection and in autoimmune disease. It is produced by the CD4 + T cells, in particular the Th17 cells. There are 6 subtypes of IL-17, from IL-17A to IL17-F, these subtypes have nearly identical structures. We know that the cytokines are interacting homotypically, but IL-17A and IL-17F are capable do perform heterotypic interaction too.
↑ Johansen C, Usher PA, Kjellerup RB, Lundsgaard D, Iversen L, Kragballe K (February 2009). "Characterization of the interleukin-17 isoforms and receptors in lesional psoriatic skin". The British Journal of Dermatology. 160 (2): 319–24. doi:10.1111/j.1365-2133.2008.08902.x. PMID19016708. S2CID205257996.
↑ Miossec P, Korn T, Kuchroo VK (August 2009). "Interleukin-17 and type 17 helper T cells". The New England Journal of Medicine. 361 (9): 888–98. doi:10.1056/NEJMra0707449. PMID19710487.
↑ Zhu H, Wang Z, Yu J, Yang X, He F, Liu Z, etal. (July 2019). "Role and mechanisms of cytokines in the secondary brain injury after intracerebral hemorrhage". Progress in Neurobiology. 178: 101610. doi:10.1016/j.pneurobio.2019.03.003. PMID30923023. S2CID85495400.
↑ Cleve H, Kirk RL, Gajdusek DC, Guiart J (1967). "On the distribution of the Gc variant Gc Aborigine in Melanesian populations; determination of Gc-types in sera from Tongariki Island, New Hebrides". Acta Genetica et Statistica Medica. 17 (6): 511–7. doi:10.1159/000152104. PMID4168861.
↑ Seppälä M, Rönnberg L, Karonen SL, Kauppila A (August 1987). "Micronized oral progesterone increases the circulating level of endometrial secretory PP14/beta-lactoglobulin homologue". Human Reproduction. 2 (6): 453–5. doi:10.1093/oxfordjournals.humrep.a136569. PMID3312283.
↑ Leonardi CL, Kimball AB, Papp KA, Yeilding N, Guzzo C, Wang Y, etal. (May 2008). "Efficacy and safety of ustekinumab, a human interleukin-12/23 monoclonal antibody, in patients with psoriasis: 76-week results from a randomised, double-blind, placebo-controlled trial (PHOENIX 1)". Lancet. 371 (9625): 1665–74. doi:10.1016/S0140-6736(08)60725-4. PMID18486739. S2CID24055733.
↑ Swardfager W, Winer DA, Herrmann N, Winer S, Lanctôt KL (March 2013). "Interleukin-17 in post-stroke neurodegeneration". Neuroscience and Biobehavioral Reviews. 37 (3): 436–47. doi:10.1016/j.neubiorev.2013.01.021. PMID23370232. S2CID10722525.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.