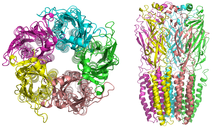
An acetylcholine receptor or a cholinergic receptor is an integral membrane protein that responds to the binding of acetylcholine, a neurotransmitter.
Muscarinic acetylcholine receptors, or mAChRs, are acetylcholine receptors that form G protein-coupled receptor complexes in the cell membranes of certain neurons and other cells. They play several roles, including acting as the main end-receptor stimulated by acetylcholine released from postganglionic fibers in the parasympathetic nervous system.

Epibatidine is a chlorinated alkaloid that is secreted by the Ecuadoran frog Epipedobates anthonyi and poison dart frogs from the Ameerega genus. It was discovered by John W. Daly in 1974, but its structure was not fully elucidated until 1992. Whether epibatidine is the first observed example of a chlorinated alkaloid remains controversial, due to challenges in conclusively identifying the compound from the limited samples collected by Daly. By the time that high-resolution spectrometry was used in 1991, there remained less than one milligram of extract from Daly's samples, raising concerns about possible contamination. Samples from other batches of the same species of frog failed to yield epibatidine.
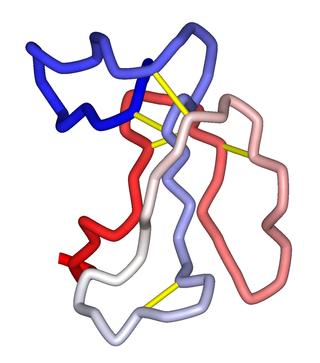
α-Bungarotoxin is one of the bungarotoxins, components of the venom of the elapid Taiwanese banded krait snake. It is a type of α-neurotoxin, a neurotoxic protein that is known to bind competitively and in a relatively irreversible manner to the nicotinic acetylcholine receptor found at the neuromuscular junction, causing paralysis, respiratory failure, and death in the victim. It has also been shown to play an antagonistic role in the binding of the α7 nicotinic acetylcholine receptor in the brain, and as such has numerous applications in neuroscience research.
Autosomal dominant nocturnal frontal lobe epilepsy is an epileptic disorder that causes frequent violent seizures during sleep. These seizures often involve complex motor movements, such as hand clenching, arm raising/lowering, and knee bending. Vocalizations such as shouting, moaning, or crying are also common. ADNFLE is often misdiagnosed as nightmares. Attacks often occur in clusters and typically first manifest in childhood. There are four known loci for ADNFLE, three with known causative genes. These genes, CHRNA4, CHRNB2, and CHRNA2, encode various nicotinic acetylcholine receptor α and β subunits.

Methyllycaconitine (MLA) is a diterpenoid alkaloid found in many species of Delphinium (larkspurs). In common with many other diterpenoid alkaloids, it is toxic to animals, although the acute toxicity varies with species. Early research was focused on identifying, and characterizing the properties of methyllycaconitine as one of the principal toxins in larkspurs responsible for livestock poisoning in the mountain rangelands of North America. Methyllycaconitine has been explored as a possible therapeutic agent for the treatment of spastic paralysis, and it has been shown to have insecticidal properties. Most recently, it has become an important molecular probe for studying the pharmacology of the nicotinic acetylcholine receptor.
A nicotinic agonist is a drug that mimics the action of acetylcholine (ACh) at nicotinic acetylcholine receptors (nAChRs). The nAChR is named for its affinity for nicotine.

α-Cobratoxin is a substance of the venom of certain Naja cobras. It is a nicotinic acetylcholine receptor (nAChR) antagonist which causes paralysis by preventing the binding of acetylcholine to the nAChR.

Neuronal acetylcholine receptor subunit alpha-4, also known as nAChRα4, is a protein that in humans is encoded by the CHRNA4 gene. The protein encoded by this gene is a subunit of certain nicotinic acetylcholine receptors (nAChR). Alpha4-containing nAChRs appear to play a crucial role in the addictive response to nicotine.

Neuronal acetylcholine receptor subunit beta-2 is a protein that in humans is encoded by the CHRNB2 gene.

Neuronal acetylcholine receptor subunit alpha-3, also known as nAChRα3, is a protein that in humans is encoded by the CHRNA3 gene. The protein encoded by this gene is a subunit of certain nicotinic acetylcholine receptors (nAchR). Research with mecamylamine in animals has implicated alpha-3-containing nAChRs in the abusive and addictive properties of ethanol.

Neuronal acetylcholine receptor subunit alpha-1, also known as nAChRα1, is a protein that in humans is encoded by the CHRNA1 gene. The protein encoded by this gene is a subunit of certain nicotinic acetylcholine receptors (nAchR).
The alpha-4 beta-2 nicotinic receptor, also known as the α4β2 receptor, is a type of nicotinic acetylcholine receptor implicated in learning, consisting of α4 and β2 subunits. It is located in the brain, where activation yields post- and presynaptic excitation, mainly by increased Na+ and K+ permeability.
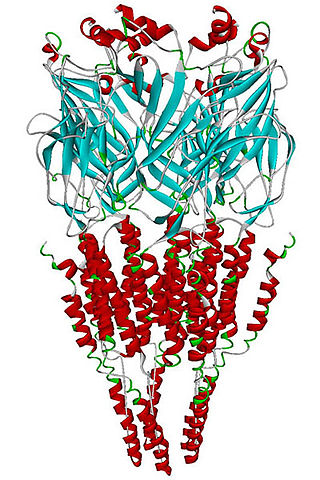
The alpha-7 nicotinic receptor, also known as the α7 receptor, is a type of nicotinic acetylcholine receptor implicated in long-term memory, consisting entirely of α7 subunits. As with other nicotinic acetylcholine receptors, functional α7 receptors are pentameric [i.e., (α7)5 stoichiometry].

Neuronal acetylcholine receptor subunit alpha-2, also known as nAChRα2, is a protein that in humans is encoded by the CHRNA2 gene. The protein encoded by this gene is a subunit of certain nicotinic acetylcholine receptors (nAchR).
Cholinergic receptor, nicotinic, alpha 6, also known as nAChRα6, is a protein that in humans is encoded by the CHRNA6 gene. The CHRNA6 gene codes for the α6 nicotinic receptor subunit that is found in certain types of nicotinic acetylcholine receptors found primarily in the brain. Neural nicotinic acetylcholine receptors containing α6 subunits are expressed on dopamine-releasing neurons in the midbrain, and dopamine release following activation of these neurons is thought to be involved in the addictive properties of nicotine. Due to their selective localisation on dopaminergic neurons, α6-containing nACh receptors have also been suggested as a possible therapeutic target for the treatment of Parkinson's disease. In addition to nicotine, research in animals has implicated alpha-6-containing nAChRs in the abusive and addictive properties of ethanol, with mecamylamine demonstrating a potent ability to block these properties.
The alpha-3 beta-4 nicotinic receptor, also known as the α3β4 receptor and the ganglion-type nicotinic receptor, is a type of nicotinic acetylcholine receptor, consisting of α3 and β4 subunits. It is located in the autonomic ganglia and adrenal medulla, where activation yields post- and/or presynaptic excitation, mainly by increased Na+ and K+ permeability.
The alpha-3 beta-2 nicotinic receptor, also known as the α3β2 receptor, is a type of nicotinic acetylcholine receptor, consisting of α3 and β2 subunits.
Anabaseine (3,4,5,6-tetrahydro-2,3′-bipyridine) is an alkaloid toxin produced by Nemertines and Aphaenogaster ants. It is structurally similar to nicotine and anabasine. Similarly, it has been shown to act as an agonist on most nicotinic acetylcholine receptors in the central nervous system and peripheral nervous system.
The neuronal acetylcholine receptor subunit alpha-5, or alpha-5 nicotinic acetylcholine receptor(α5 nAChR) also known as the α5 receptor is a type of ligand gated neuronal type subunit of the nicotinic acetylcholine receptor involved in pain regulation encoded in the human by the CHRNA5 gene. This receptor is commonly associated with nicotine addiction, immunotherapy, cancer, pain and attention.