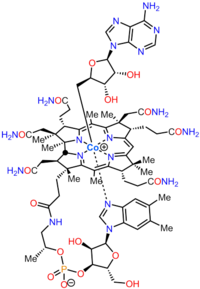
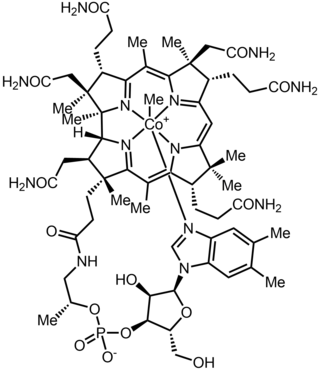
Last updated AdenosylcobalaminMethylcobalamin, another biologically active form. The dark red crystals dissolve in water giving cherry-colored solutions.
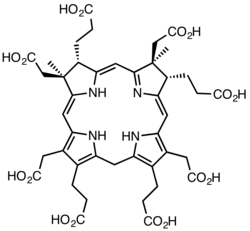
Cobalamin biosynthesis is the process by which bacteria and archea make cobalamin, vitamin B12. Many steps are involved in converting aminolevulinic acid via uroporphyrinogen III and adenosylcobyric acid to the final forms in which it is used by enzymes in both the producing organisms and other species, including humans who acquire it through their diet.
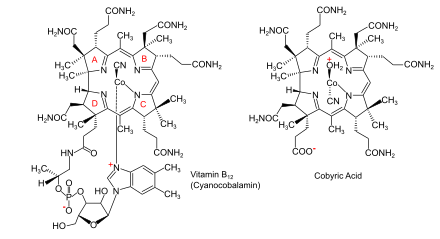
The feature which distinguishes the two main biosynthetic routes is whether the cobalt that is at the catalytic site in the coenzyme is incorporated early (in anaerobic organisms) or late (in aerobic organisms) and whether oxygen is required. In both cases, the macrocycle that will form a coordination complex with the cobalt ion is a corrin ring, specifically one with seven carboxylate groups called cobyrinic acid. Subsequently, amide groups are formed on all but one of the carboxylates, giving cobyric acid, and the cobalt is ligated by an adenosyl group. In the final part of the biosynthesis, common to all organisms, an aminopropanol sidechain is added to the one free carboxylic group and assembly of the nucleotide loop, which will provide the second ligand for the cobalt, is completed.
Many prokaryotic species cannot biosynthesize adenosylcobalamin, but can make it from cobalamin which they assimilate from external sources. In humans, dietary sources of cobalamin are bound after ingestion as transcobalamins and converted to the coenzyme forms in which they are used.
Corrin ring synthesis leading to cobyrinic acid, with seven carboxylate groups. In the anaerobic pathway this already contains cobalt but in the aerobic pathway the material formed at that stage is hydrogenobyrinic acid, without the bound cobalt.[11][12][5]
Insertion of cobalt, where not already present; formation of amides on all but one of the carboxylate groups to give cobyric acid; attachment of an adenosyl group as ligand to the cobalt; attachment of an aminopropanol sidechain to the one free carboxylic group and assembly of the nucleotide loop which will provide the second ligand for the cobalt.[5][13]
Detail of steps from uroporphyrinogen III to cob(II)yrinic acid a,c-diamide in aerobic organisms
The biosynthesis of cobalamin diverges from that of haem and chlorophyll at uroporphrinogen III: its transformation involves the sequential addition of methyl (CH3) groups to give intermediates that were given trivial names according to the number of these groups that have been incorporated. Hence, the first intermediate is precorrin-1, the next is precorrin-2 and so on. The incorporation of all eight additional methyl groups which occur in cobyric acid was investigated using 13C methyl-labelledS-adenosyl methionine. It was not until scientists at Rhône-Poulenc Rorer used a genetically-engineered strain of Pseudomonas denitrificans, in which eight of the cob genes involved in the biosynthesis of the vitamin had been overexpressed, that the complete sequence of methylation and other steps could be determined, thus fully establishing all the intermediates in the pathway.[18][19]
This enzyme is an oxidoreductase that requires oxygen and hence the reaction can only operate under aerobic conditions. The naming of these precorrins as 3A and 3B reflects the fact that each contains three more methyl groups than uroporphyrinogen III but with different structures: in particular, precorrin-3B has an internal γ-lactone ring formed from the ring A acetic acid sidechain closing back on to the macrocycle.
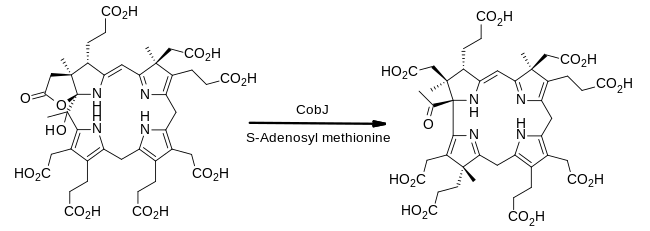
From precorrin-3B to precorrin-4
The enzyme CobJ continues the theme of methyl group insertion. Importantly, during this step the macrocyclering-contracts so that the product contains for the first time the corrin core which characterises cobalamin.[18]
The newly-inserted methyl group is added to ring C at the carbon attached to the methylene (CH2) bridge to ring B. This is not its final location on cobalamin as a later step involves its rearrangement to an adjacent ring carbon.
From precorrin-5 to precorrin-6A
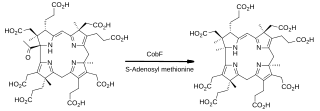
The enzyme CobF now removes the acetyl group located at position 1 of the ring system in precorrin-4 and replaces it with a newly-introduced methyl group. The name of the product, precorrin-6A, reflects the fact that six methyl groups in total have been added to uroporphyrinogen III up to this point. However, since one of these has been extruded with the acetate group, the structure of precorrin-6A contains just the remaining five.[21]
Precorrin-6B therefore differs in structure from precorrin-6A only by having an extra two hydrogen atoms.
From precorrin-6B to precorrin-8
The enzyme CobL has two active sites, one catalysing two methyl group additions and the other the decarboxylation of the CH2COOH group on ring D, so that this substituent becomes a simple methyl group:[21]
The enzyme CobH catalyzes a rearrangement reaction, with the result that the methyl group that had been added to ring C is isomerised to its final location, an example of intramolecular transfer:[22]
precorrin-8X hydrogenobyrinate
From hydrogenobyrinic acid to hydrogenobyrinic acid a,c-diamide
The next enzyme in the pathway, CobB, selectively converts two of the eight carboxylic acid groups into their primary amides. ATP is used to provide the energy for amide bond formation, with the transferred ammonia coming from glutamine:[23]
It is at this stage that the aerobic pathway and the anaerobic pathway merge, with later steps being chemically identical.
Detail of steps from uroporphyrinogen III to cob(II)yrinic acid a,c-diamide in anaerobic organisms
Many of the steps beyond uroporphyrinogen III in anaerobic organisms such as Bacillus megaterium involve chemically similar but genetically distinct transformations to those in the aerobic pathway.[10][25]
As in the aerobic pathway, the third methyl group is introduced by a methyltransferase enzyme, CbiL:[26]
cobalt-sirohydrochlorin + S-adenosyl methionine cobalt-factor III + S-adenosyl-L-homocysteine
From cobalt-factor III to cobalt-precorrin-4
Methylation and ring contraction to form the corrin macrocycle occurs next, catalysed by the enzyme Cobalt-factor III methyltransferase (CbiH, EC2.1.1.272)[28]
cobalt-factor III + S-adenosyl methionine cobalt-precorrin-4 + S-adenosyl-L-homocysteine
In this pathway, the resulting material contains a δ-lactone, a six-membered ring, rather than the γ-lactone (five-membered ring) of precorrin-3B.
From cobalt-precorrin-4 to cobalt-precorrin-5A
The introduction of the methyl group at C-11 in the next step is catalysed by Cobalt-precorrin-4 methyltransferase (CbiF, EC2.1.1.271)[29]
The scene is now set for the extrusion of the two-carbon fragment corresponding to the acetate released in the formation of precorrin-6A in the aerobic pathway. In this case the fragment released is acetaldehyde and this is catalysed by CbiG:[29]
From cobalt-precorrin-5B to cob(II)yrinic acid a,c-diamide
The steps from cobalt-precorrin-5B to cob(II)yrinic acid a,c-diamide in the anaerobic pathway are essentially chemically identical to those in the aerobic sequence. The intermediates are called cobalt-precorrin-6A, cobalt-precorrin-6B, cobalt-precorrin-8 and cobyrinic acid. The enzymes in sequence are CbiD;[30] Cobalt-precorrin-6A reductase (CbiJ, EC1.3.1.106);[31]CbiT, Cobalt-precorrin-8 methylmutase (CbiC, EC5.4.99.60) and CbiA. The final enzyme forms cob(II)yrinic acid a,c-diamide as the two pathways converge.[5]
Detail of steps from cob(II)yrinic acid a,c-diamide to adenosylcobalamin
Aerobic and anaerobic organisms share the same chemical pathway beyond cob(II)yrinic acid a,c-diamide and this is illustrated for the cob gene products.
From cob(II)yrinic acid a,c-diamide to adenosylcobyric acid
The cobalt(II) is reduced to cobalt(I) by the enzyme CobR and then the enzyme CobO attaches an adenosyl ligand to the metal. Next, the enzyme CobQ converts all the carboxylic acids, except the propionic acid on ring D, to their primary amides.[7][21]
From adenosylcobyric acid to adenosylcobinamide phosphate
In aerobic organisms, the enzyme CobCD now attaches (R)-1-amino-2-propanol (derived from threonine) to the propionic acid, forming adenosylcobinamide and the enzyme CobUphosphorylates the terminal hydroxy group to form adenosylcobinamide phosphate.[21] The same final product is formed in anaerobic organisms by direct reaction of adenosylcobyric acid with (R)-1-amino-2-propanol O-2-phosphate (derived from threonine-O-phosphate by the enzyme CobD) catalysed by the enzyme CbiB.[5]
From adenosylcobinamide phosphate to adenosylcobalamin
In a separate branch of the pathway, 5,6-dimethylbenzimidazole is biosynthesised from flavin mononucleotide by the enzyme 5,6-dimethylbenzimidazole synthase and converted by CobT to alpha-ribazole 5' phosphate. Then the enzyme CobU activates adenosylcobinamide phosphate by formation of adenosylcobinamide-GDP and CobV links the two substrates to form Adenosylcobalamin-5'-phosphate. In the final step to the coenzyme, CobC removes the 5' phosphate group:[32][33]
The complete biosynthetic route involves a long linear path that requires about 25 contributing enzyme steps.
Other pathways of cobalamin metabolism
Salvage pathways in prokaryotes
Many prokaryotic species cannot biosynthesize adenosylcobalamin, but can make it from cobalamin. These organisms are capable of cobalamin transport into the cell and its conversion to the required coenzyme form.[34] Even organisms such as Salmonella typhimurium that can make cobalamin also assimilate it from external sources when available.[5][35][36][37] Uptake into cells is facilitated by ABC transporters which absorb the cobalamin through the cell membrane.[38]
Methylcobalamin (mecobalamin, MeCbl, or MeB12) is a cobalamin, a form of vitamin B12. It differs from cyanocobalamin in that the cyano group at the cobalt is replaced with a methyl group. Methylcobalamin features an octahedral cobalt(III) centre and can be obtained as bright red crystals. From the perspective of coordination chemistry, methylcobalamin is notable as a rare example of a compound that contains metal–alkyl bonds. Nickel–methyl intermediates have been proposed for the final step of methanogenesis.
In enzymology, a cobalt-factor II C20-methyltransferase is an enzyme that catalyzes the chemical reaction
In enzymology, a precorrin-2 C20-methyltransferase is an enzyme that catalyzes the chemical reaction
In enzymology, precorrin-3B C17-methyltransferase is an enzyme that catalyzes the chemical reaction
In enzymology, a precorrin-4 C11-methyltransferase is an enzyme that catalyzes the chemical reaction
In enzymology, precorrin-6A synthase (deacetylating) (EC 2.1.1.152) is an enzyme that catalyzes the chemical reaction
In enzymology, a precorrin-6Y C5,15-methyltransferase (decarboxylating) (EC 2.1.1.132) is an enzyme that catalyzes the chemical reaction
In enzymology, a precorrin-6A reductase (EC 1.3.1.54) is an enzyme that catalyzes the chemical reaction
In enzymology, a precorrin-3B synthase (EC 1.14.13.83) is an enzyme that catalyzes the chemical reaction
In enzymology, a precorrin-8X methylmutase is an enzyme that catalyzes the chemical reaction
Cobalt chelatase (EC 6.6.1.2) is an enzyme that catalyzes the chemical reaction
In enzymology, a hydrogenobyrinic acid a,c-diamide synthase (glutamine-hydrolysing) (EC 6.3.5.9) is an enzyme that catalyzes the chemical reaction
The primary biochemical reaction catalyzed by the enzyme adenosylcobalamin/α-ribazole phosphatase (formerly α-ribazole phosphatase) (EC 3.1.3.73) is
In enzymology, a nicotinate-nucleotide-dimethylbenzimidazole phosphoribosyltransferase is an enzyme that catalyzes the chemical reaction
In molecular biology, the vitamin B12-binding domain is a protein domain which binds to cobalamin. It can bind two different forms of the cobalamin cofactor, with cobalt bonded either to a methyl group (methylcobalamin) or to 5'-deoxyadenosine (adenosylcobalamin). Cobalamin-binding domains are mainly found in two families of enzymes present in animals and prokaryotes, which perform distinct kinds of reactions at the cobalt-carbon bond. Enzymes that require methylcobalamin carry out methyl transfer reactions. Enzymes that require adenosylcobalamin catalyse reactions in which the first step is the cleavage of adenosylcobalamin to form cob(II)alamin and the 5'-deoxyadenosyl radical, and thus act as radical generators. In both types of enzymes the B12-binding domain uses a histidine to bind the cobalt atom of cobalamin cofactors. This histidine is embedded in a DXHXXG sequence, the most conserved primary sequence motif of the domain. Proteins containing the cobalamin-binding domain include:
Uroporphyrinogen-III C-methyltransferase, uroporphyrinogen methyltransferase, uroporphyrinogen-III methyltransferase, adenosylmethionine-uroporphyrinogen III methyltransferase, S-adenosyl-L-methionine-dependent uroporphyrinogen III methylase, uroporphyrinogen-III methylase, SirA, CysG, CobA, uroporphyrin-III C-methyltransferase, S-adenosyl-L-methionine:uroporphyrin-III C-methyltransferase) is an enzyme with systematic name S-adenosyl-L-methionine:uroporphyrinogen-III C-methyltransferase. This enzyme catalyses the following chemical reaction
Cobalt-precorrin-5B (C1)-methyltransferase (EC 2.1.1.195), cobalt-precorrin-6A synthase, CbiD (gene)) is an enzyme with systematic name S-adenosyl-L-methionine:cobalt-precorrin-5B (C1)-methyltransferase. This enzyme catalyses the following chemical reaction
Cobalt-precorrin-7 (C15)-methyltransferase (decarboxylating) (EC 2.1.1.196, CbiT) is an enzyme with systematic name S-adenosyl-L-methionine:precorrin-7 C15-methyltransferase (C12-decarboxylating). This enzyme catalyses the following chemical reaction
Adenosylcobinamide-phosphate synthase is an enzyme with systematic name adenosylcobyric acid:(R)-1-aminopropan-2-yl phosphate ligase (ADP-forming). This enzyme catalyses the following chemical reaction
Cobyrinate a,c-diamide synthase (EC ), cobyrinic acid a,c-diamide synthetase, CbiA (gene)) is an enzyme which catalyses the chemical reaction
↑ Banerjee R (2006). "B12 Trafficking in Mammals: A Case for Coenzyme Escort Service". ACS Chemical Biology. 1 (3): 149–159. doi:10.1021/cb6001174. PMID17163662.
↑ "Vitamin B12". Micronutrient Information Center, Linus Pauling Institute, Oregon State University, Corvallis, OR. 4 June 2015. Retrieved 20 April 2020.
↑ Roessner CA, Santander PJ, Scott A (2001). "Multiple biosynthetic pathways for vitamin B12: Variations on a central theme". Cofactor Biosynthesis. Vitamins & Hormones. Vol.61. pp.267–297. doi:10.1016/s0083-6729(01)61009-4. ISBN9780127098616. PMID11153269.
↑ Heldt D, Lawrence A, Lindenmeyer M, Deery E, Heathcote P, Rigby S, Warren M (2005). "Aerobic synthesis of vitamin B12: Ring contraction and cobalt chelation". Biochemical Society Transactions. 33 (4): 815–819. doi:10.1042/BST0330815. PMID16042605. S2CID37362827.
↑ Frank S, Brindley A, Deery E, Heathcote P, Lawrence A, Leech H, Pickersgill R, Warren M (2005). "Anaerobic synthesis of vitamin B12: Characterization of the early steps in the pathway". Biochemical Society Transactions. 33 (4): 811–814. doi:10.1042/BST0330811. PMID16042604.
↑ Battersby AR (2000). "Tetrapyrroles: the Pigments of Life. A Millennium review". Nat. Prod. Rep. 17 (6): 507–526. doi:10.1039/B002635M. PMID11152419.
↑ Raux E, Schubert HL, Warren MJ (December 2000). "Biosynthesis of cobalamin (vitamin B12): a bacterial conundrum". Cell. Mol. Life Sci. 57 (13–14): 1880–1893. doi:10.1007/PL00000670. PMID11215515. S2CID583311.
↑ Frank S, Brindley AA, Deery E, Heathcote P, Lawrence AD, Leech HK, etal. (August 2005). "Anaerobic synthesis of vitamin B12: characterization of the early steps in the pathway". Biochemical Society Transactions. 33 (Pt 4): 811–814. doi:10.1042/BST0330811. PMID16042604.
↑ Battersby A (2005). "Chapter 11: Discovering the wonder of how Nature builds its molecules". In Archer MD, Haley CD (eds.). The 1702 chair of chemistry at Cambridge: transformation and change. Cambridge University Press. pp.xvi, 257–282. ISBN0521828732.
1 2 Kajiwara Y, Santander PJ, Roessner CA, Pérez LM, Scott AI (2006). "Genetically Engineered Synthesis and Structural Characterization of Cobalt−Precorrin 5A and −5B, Two New Intermediates on the Anaerobic Pathway to Vitamin B12: Definition of the Roles of the CbiF and CbiG Enzymes". Journal of the American Chemical Society. 128 (30): 9971–9978. doi:10.1021/ja062940a. PMID16866557.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.