Related Research Articles

In molecular biology, RNA polymerase, or more specifically DNA-directed/dependent RNA polymerase (DdRP), is an enzyme that catalyzes the chemical reactions that synthesize RNA from a DNA template.

Chromista is a proposed but polyphyletic biological kingdom consisting of single-celled and multicellular eukaryotic species that share similar features in their photosynthetic organelles (plastids). It includes all eukaryotes whose plastids contain chlorophyll c and are surrounded by four membranes. If the ancestor already possessed chloroplasts derived by endosymbiosis from red algae, all non-photosynthetic Chromista have secondarily lost the ability to photosynthesise. Its members might have arisen independently as separate evolutionary groups from the last eukaryotic common ancestor.

Viridiplantae constitute a clade of eukaryotic organisms that comprises approximately 450,000–500,000 species that play important roles in both terrestrial and aquatic ecosystems. They include the green algae, which are primarily aquatic, and the land plants (embryophytes), which emerged from within them. Green algae traditionally excludes the land plants, rendering them a paraphyletic group. However it is accurate to think of land plants as a kind of alga. Since the realization that the embryophytes emerged from within the green algae, some authors are starting to include them. They have cells with cellulose in their cell walls, and primary chloroplasts derived from endosymbiosis with cyanobacteria that contain chlorophylls a and b and lack phycobilins. Corroborating this, a basal phagotroph archaeplastida group has been found in the Rhodelphydia.
A release factor is a protein that allows for the termination of translation by recognizing the termination codon or stop codon in an mRNA sequence. They are named so because they release new peptides from the ribosome.

Elongation factors are a set of proteins that function at the ribosome, during protein synthesis, to facilitate translational elongation from the formation of the first to the last peptide bond of a growing polypeptide. Most common elongation factors in prokaryotes are EF-Tu, EF-Ts, EF-G. Bacteria and eukaryotes use elongation factors that are largely homologous to each other, but with distinct structures and different research nomenclatures.
EF-Tu is a prokaryotic elongation factor responsible for catalyzing the binding of an aminoacyl-tRNA (aa-tRNA) to the ribosome. It is a G-protein, and facilitates the selection and binding of an aa-tRNA to the A-site of the ribosome. As a reflection of its crucial role in translation, EF-Tu is one of the most abundant and highly conserved proteins in prokaryotes. It is found in eukaryotic mitochondria as TUFM.
Eukaryotic initiation factors (eIFs) are proteins or protein complexes involved in the initiation phase of eukaryotic translation. These proteins help stabilize the formation of ribosomal preinitiation complexes around the start codon and are an important input for post-transcription gene regulation. Several initiation factors form a complex with the small 40S ribosomal subunit and Met-tRNAiMet called the 43S preinitiation complex. Additional factors of the eIF4F complex recruit the 43S PIC to the five-prime cap structure of the mRNA, from which the 43S particle scans 5'-->3' along the mRNA to reach an AUG start codon. Recognition of the start codon by the Met-tRNAiMet promotes gated phosphate and eIF1 release to form the 48S preinitiation complex, followed by large 60S ribosomal subunit recruitment to form the 80S ribosome. There exist many more eukaryotic initiation factors than prokaryotic initiation factors, reflecting the greater biological complexity of eukaryotic translation. There are at least twelve eukaryotic initiation factors, composed of many more polypeptides, and these are described below.
Elongation factor 4 (EF-4) is an elongation factor that is thought to back-translocate on the ribosome during the translation of RNA to proteins. It is found near-universally in bacteria and in eukaryotic endosymbiotic organelles including the mitochondria and the plastid. Responsible for proofreading during protein synthesis, EF-4 is a recent addition to the nomenclature of bacterial elongation factors.
Protein-synthesizing GTPases are enzymes involved in mRNA translation into protein by the ribosome, with systematic name GTP phosphohydrolase (mRNA-translation-assisting). They usually include translation initiation factors such as IF-2 and translation elongation factors such as EF-Tu.

Elongation factor 1-alpha 1 (eEF1a1) is a translation elongation protein, expressed across eukaryotes. In humans, it is encoded by the EEF1A1 gene.

Eukaryotic elongation factor 2 is a protein that in humans is encoded by the EEF2 gene. It is the archaeal and eukaryotic counterpart of bacterial EF-G.

EF-G is a prokaryotic elongation factor involved in protein translation. As a GTPase, EF-G catalyzes the movement (translocation) of transfer RNA (tRNA) and messenger RNA (mRNA) through the ribosome.
EF-Ts is one of the prokaryotic elongation factors. It is found in human mitochondria as TSFM. It is similar to eukaryotic EF-1B.
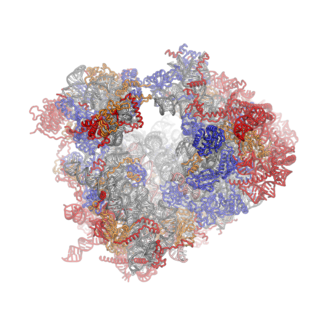
Ribosomes are a large and complex molecular machine that catalyzes the synthesis of proteins, referred to as translation. The ribosome selects aminoacylated transfer RNAs (tRNAs) based on the sequence of a protein-encoding messenger RNA (mRNA) and covalently links the amino acids into a polypeptide chain. Ribosomes from all organisms share a highly conserved catalytic center. However, the ribosomes of eukaryotes are much larger than prokaryotic ribosomes and subject to more complex regulation and biogenesis pathways. Eukaryotic ribosomes are also known as 80S ribosomes, referring to their sedimentation coefficients in Svedberg units, because they sediment faster than the prokaryotic (70S) ribosomes. Eukaryotic ribosomes have two unequal subunits, designated small subunit (40S) and large subunit (60S) according to their sedimentation coefficients. Both subunits contain dozens of ribosomal proteins arranged on a scaffold composed of ribosomal RNA (rRNA). The small subunit monitors the complementarity between tRNA anticodon and mRNA, while the large subunit catalyzes peptide bond formation.

In molecular biology, the EF1 guanine nucleotide exchange domain is a protein domain found in the beta and delta chains of elongation factors from eukaryotes and archaea.

The eocyte hypothesis in evolutionary biology proposes that the eukaryotes originated from a group of prokaryotes called eocytes. After his team at the University of California, Los Angeles discovered eocytes in 1984, James A. Lake formulated the hypothesis as "eocyte tree" that proposed eukaryotes as part of archaea. Lake hypothesised the tree of life as having only two primary branches: prokaryotes, which include Bacteria and Archaea, and karyotes, that comprise Eukaryotes and eocytes. Parts of this early hypothesis were revived in a newer two-domain system of biological classification which named the primary domains as Archaea and Bacteria.

Archaeal transcription factor B is a protein family of extrinsic transcription factors that guide the initiation of RNA transcription in organisms that fall under the domain of Archaea. It is homologous to eukaryotic TFIIB and, more distantly, to bacterial sigma factor. Like these proteins, it is involved in forming transcription preinitiation complexes. Its structure includes several conserved motifs which interact with DNA and other transcription factors, notably the single type of RNA polymerase that performs transcription in Archaea.
Lokiarchaeota is a proposed phylum of the Archaea. The phylum includes all members of the group previously named Deep Sea Archaeal Group, also known as Marine Benthic Group B. Lokiarchaeota is part of the superphylum Asgard containing the phyla: Lokiarchaeota, Thorarchaeota, Odinarchaeota, Heimdallarchaeota, and Helarchaeota. A phylogenetic analysis disclosed a monophyletic grouping of the Lokiarchaeota with the eukaryotes. The analysis revealed several genes with cell membrane-related functions. The presence of such genes support the hypothesis of an archaeal host for the emergence of the eukaryotes; the eocyte-like scenarios.
Archaeal initiation factors are proteins that are used during the translation step of protein synthesis in archaea. The principal functions these proteins perform include ribosome RNA/mRNA recognition, delivery of the initiator Met-tRNAiMet, methionine bound tRNAi, to the 40s ribosome, and proofreading of the initiation complex.

Archaeal transcription is the process in which a segment of archaeal DNA is copied into a newly synthesized strand of RNA using the sole Pol II-like RNA polymerase (RNAP). The process occurs in three main steps: initiation, elongation, and termination; and the end result is a strand of RNA that is complementary to a single strand of DNA. A number of transcription factors govern this process with homologs in both bacteria and eukaryotes, with the core machinery more similar to eukaryotic transcription.
References
- 1 2 Andersen GR, Nyborg J (2001). "Structural studies of eukaryotic elongation factors". Cold Spring Harbor Symposia on Quantitative Biology. 66: 425–37. doi:10.1101/sqb.2001.66.425. PMID 12762045.
- ↑ Vitagliano L, Masullo M, Sica F, Zagari A, Bocchini V (October 2001). "The crystal structure of Sulfolobus solfataricus elongation factor 1alpha in complex with GDP reveals novel features in nucleotide binding and exchange". The EMBO Journal. 20 (19): 5305–11. doi: 10.1093/emboj/20.19.5305 . PMC 125647 . PMID 11574461.
- 1 2 3 4 5 Sasikumar AN, Perez WB, Kinzy TG (2011). "The many roles of the eukaryotic elongation factor 1 complex". Wiley Interdisciplinary Reviews: RNA. 3 (4): 543–55. doi:10.1002/wrna.1118. PMC 3374885 . PMID 22555874.
- ↑ Kaminska M, Havrylenko S, Decottignies P, Gillet S, Le Maréchal P, Negrutskii B, Mirande M (March 2009). "Dissection of the structural organization of the aminoacyl-tRNA synthetase complex". The Journal of Biological Chemistry. 284 (10): 6053–60. doi: 10.1074/jbc.M809636200 . PMID 19131329.
- ↑ Ellen Cocquyt; Heroen Verbruggen; Frederik Leliaert; Frederick W Zechman; Koen Sabbe; Olivier De Clerck (2009), "Gain and loss of elongation factor genes in green algae", BMC Evol. Biol., 9 (1): 39, Bibcode:2009BMCEE...9...39C, doi: 10.1186/1471-2148-9-39 , PMC 2652445 , PMID 19216746
- ↑ Kamikawa R, Brown MW, Nishimura Y, Sako Y, Heiss AA, Yubuki N, et al. (June 2013). "Parallel re-modeling of EF-1α function: divergent EF-1α genes co-occur with EFL genes in diverse distantly related eukaryotes". BMC Evolutionary Biology. 13 (1): 131. Bibcode:2013BMCEE..13..131K. doi: 10.1186/1471-2148-13-131 . PMC 3699394 . PMID 23800323.
- ↑ Mikhailov KV, Janouškovec J, Tikhonenkov DV, Mirzaeva GS, Diakin AY, Simdyanov TG, et al. (September 2014). "A complex distribution of elongation family GTPases EF1A and EFL in basal alveolate lineages". Genome Biology and Evolution. 6 (9): 2361–7. doi: 10.1093/gbe/evu186 . PMC 4217694 . PMID 25179686.
- ↑ Saito K, Kobayashi K, Wada M, Kikuno I, Takusagawa A, Mochizuki M, et al. (November 2010). "Omnipotent role of archaeal elongation factor 1 alpha (EF1α in translational elongation and termination, and quality control of protein synthesis". Proceedings of the National Academy of Sciences of the United States of America. 107 (45): 19242–7. Bibcode:2010PNAS..10719242S. doi: 10.1073/pnas.1009599107 . PMC 2984191 . PMID 20974926.