
An endosymbiont or endobiont is an organism that lives within the body or cells of another organism. Typically the two organisms are in a mutualistic relationship. Examples are nitrogen-fixing bacteria, which live in the root nodules of legumes, single-cell algae inside reef-building corals, and bacterial endosymbionts that provide essential nutrients to insects.

Aphids are small sap-sucking insects and members of the superfamily Aphidoidea. Common names include greenfly and blackfly, although individuals within a species can vary widely in color. The group includes the fluffy white woolly aphids. A typical life cycle involves flightless females giving live birth to female nymphs—who may also be already pregnant, an adaptation scientists call telescoping generations—without the involvement of males. Maturing rapidly, females breed profusely so that the number of these insects multiplies quickly. Winged females may develop later in the season, allowing the insects to colonize new plants. In temperate regions, a phase of sexual reproduction occurs in the autumn, with the insects often overwintering as eggs.
A bacteriome is a specialized organ, found mainly in some insects, that hosts endosymbiotic bacteria. Bacteriomes contain specialized cells, called bacteriocytes, that provide nutrients and shelter to the bacteria while protecting the host animal. In exchange, the bacteria provide essentials like vitamins and amino acids to the host insect. Bacteriomes also protect the bacteria from the host's immune system, with insects secreting antimicrobial peptides such as the coleoptericin secreted by weevils to keep bacteria within the bacteriome.

Buchnera aphidicola, a member of the Pseudomonadota and the only species in the genus Buchnera, is the primary endosymbiont of aphids, and has been studied in the pea aphid, Acyrthosiphon pisum. Buchnera is believed to have had a free-living, Gram-negative ancestor similar to a modern Enterobacterales, such as Escherichia coli. Buchnera is 3 μm in diameter and has some of the key characteristics of its Enterobacterales relatives, such as a Gram-negative cell wall. However, unlike most other Gram-negative bacteria, Buchnera lacks the genes to produce lipopolysaccharides for its outer membrane. The long association with aphids and the limitation of crossover events due to strictly vertical transmission has seen the deletion of genes required for anaerobic respiration, the synthesis of amino sugars, fatty acids, phospholipids, and complex carbohydrates. This has resulted not only in one of the smallest known genomes of any living organism, but also one of the most genetically stable.
Wigglesworthia glossinidia is a species of gram-negative bacteria that is a bacterial endosymbiont of the tsetse fly. Because of this relationship, Wigglesworthia has lost a large part of its genome, leaving it with one of the smallest genomes of any living organism, consisting of a single chromosome of 700,000 bp and a plasmid of 5,200. Together with Buchnera aphidicola, Wigglesworthia has been the subject of genetic research into the minimal genome necessary for any living organism.

Acyrthosiphon pisum, commonly known as the pea aphid, is a sap-sucking insect in the family Aphididae. It feeds on several species of legumes worldwide, including forage crops, such as pea, clover, alfalfa, and broad bean, and ranks among the aphid species of major agronomical importance. The pea aphid is a model organism for biological study whose genome has been sequenced and annotated.
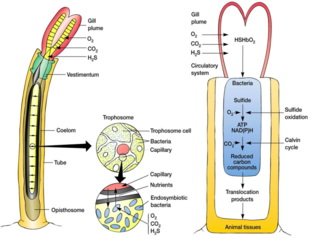
A trophosome is a highly vascularised organ found in some animals that houses symbiotic bacteria that provide food for their host. Trophosomes are contained by the coelom of tube worms and in the body of symbiotic flatworms of the genus Paracatenula.
The hologenome theory of evolution recasts the individual animal or plant as a community or a "holobiont" – the host plus all of its symbiotic microbes. Consequently, the collective genomes of the holobiont form a "hologenome". Holobionts and hologenomes are structural entities that replace misnomers in the context of host-microbiota symbioses such as superorganism, organ, and metagenome. Variation in the hologenome may encode phenotypic plasticity of the holobiont and can be subject to evolutionary changes caused by selection and drift, if portions of the hologenome are transmitted between generations with reasonable fidelity. One of the important outcomes of recasting the individual as a holobiont subject to evolutionary forces is that genetic variation in the hologenome can be brought about by changes in the host genome and also by changes in the microbiome, including new acquisitions of microbes, horizontal gene transfers, and changes in microbial abundance within hosts. Although there is a rich literature on binary host–microbe symbioses, the hologenome concept distinguishes itself by including the vast symbiotic complexity inherent in many multicellular hosts.

The minimal genome is a concept which can be defined as the set of genes sufficient for life to exist and propagate under nutrient-rich and stress-free conditions. Alternatively, it may be defined as the gene set supporting life on an axenic cell culture in rich media, and it is thought what makes up the minimal genome will depend on the environmental conditions that the organism inhabits.
Hamiltonella defensa is a species of bacteria. It is maternally or sexually transmitted and lives as an endosymbiont of whiteflies and aphids, meaning that it lives within a host, protecting its host from attack. It does this through bypassing the host's immune responses by protecting its host against parasitoid wasps. However, H. defensa is only defensive if infected by a virus. H. defensa shows a relationship with Photorhabdus species, together with Regiella insecticola. Together with other endosymbionts, it provides aphids protection against parasitoids. It is known to habitate Bemisia tabaci.

Angomonas deanei is a flagellated trypanosomatid protozoan. As an obligate parasite, it infects the gastrointestinal tract of insects, and is in turn a host to symbiotic bacteria. The bacterial endosymbiont Ca. "Kinetoplastibacterium crithidii" maintains a permanent mutualistic relationship with the protozoan such that it is no longer able to reproduce and survive on its own. The symbiosis, subsequently also discovered in varying degrees in other protists such as Strigomonas culicis, Novymonas esmeraldas, Diplonema japonicumand Diplonema aggregatum are considered as good models for the understanding of the evolution of eukaryotes from prokaryotes, and on the origin of cell organelles.
Arsenophonus nasoniae is a species of bacterium which was previously isolated from Nasonia vitripennis, a species of parasitoid wasp. These wasps are generalists which afflict the larvae of parasitic carrion flies such as blowflies, houseflies and flesh flies. A. nasoniae belongs to the phylum Pseudomonadota and family Morganellaceae. The genus Arsenophonus, has a close relationship to the Proteus (bacterium) rather than to that of Salmonella and Escherichia. The genus is composed of gammaproteobacterial, secondary-endosymbionts which are gram-negative. Cells are non-flagellated, non-motile, non-spore forming and form long to highly filamentous rods. Cellular division is exhibited through septation. The name 'Arsenophonus nasoniae gen. nov., sp. nov.' was therefore proposed for the discovered bacterium due to its characteristics and its microbial interaction with N. vitripennis. The type strain of A. nasoniae is Strain SKI4.
Nasuia deltocephalinicola was reported in 2013 to have the smallest genome of all bacteria, with 112,091 nucleotides. For comparison, the genome of Escherichia coli has 4.6 million nucleotides. The second smallest genome, from bacteria Tremblaya princeps, has 139,000 nucleotides. While N. deltocephalinicola has the smallest number of nucleotides, it has more protein-coding genes (137) than some bacteria.

Paracatenula is a genus of millimeter sized free-living marine gutless catenulid flatworms.
The initial acquisition of microbiota is the formation of an organism's microbiota immediately before and after birth. The microbiota are all the microorganisms including bacteria, archaea and fungi that colonize the organism. The microbiome is another term for microbiota or can refer to the collected genomes.
"Candidatus Karelsulcia muelleri" is an aerobic, gram-negative, bacillus bacterium that is a part of the phylum Bacteroidota. "Ca. K. muelleri" is an obligate and mutualistic symbiotic microbe commonly found occupying specialized cell compartments of sap-feeding insects called bacteriocytes. A majority of the research done on "Ca. K. muelleri" has detailed its relationship with the host Homalodisca vitripennis. Other studies have documented the nature of its residency in other insects like the maize leafhopper (Cicadulina) or the spittlebug (Cercopoidea). "Ca. K. muelleri" is noted for its exceptionally minimal genome and it is currently identified as having the smallest known sequenced Bacteroidota genome at only 245 kilobases.

A symbiosome is a specialised compartment in a host cell that houses an endosymbiont in a symbiotic relationship.
Vertical transmission of symbionts is the transfer of a microbial symbiont from the parent directly to the offspring. Many metazoan species carry symbiotic bacteria which play a mutualistic, commensal, or parasitic role. A symbiont is acquired by a host via horizontal, vertical, or mixed transmission.
Glossina fuscipes is a riverine fly species in the genus Glossina, which are commonly known as tsetse flies. Typically found in sub-Saharan Africa but with a small Arabian range, G. fuscipes is a regional vector of African trypanosomiasis, commonly known as sleeping sickness, that causes significant rates of morbidity and mortality among humans and livestock. Consequently, the species is among several being targeted by researchers for population control as a method for controlling the disease.

Strigomonas culicis is a protist and member of flagellated trypanosomatids. It is an obligate parasite in the gastrointestinal tract of mosquito, and is in turn a host to symbiotic bacteria. It maintains strict mutualistic relationship with the bacteria as a sort of cell organelle (endosymbiont) so that it cannot lead an independent life without the bacteria. Along with Angomonas deanei, S. culicis is researched as model organism for the evolution of symbiotic relationsships with intracellular bacteria.



