Related Research Articles
A germ cell is any biological cell that gives rise to the gametes of an organism that reproduces sexually. In many animals, the germ cells originate in the primitive streak and migrate via the gut of an embryo to the developing gonads. There, they undergo meiosis, followed by cellular differentiation into mature gametes, either eggs or sperm. Unlike animals, plants do not have germ cells designated in early development. Instead, germ cells can arise from somatic cells in the adult, such as the floral meristem of flowering plants.

Spermatogenesis is the process by which haploid spermatozoa develop from germ cells in the seminiferous tubules of the testis. This process starts with the mitotic division of the stem cells located close to the basement membrane of the tubules. These cells are called spermatogonial stem cells. The mitotic division of these produces two types of cells. Type A cells replenish the stem cells, and type B cells differentiate into primary spermatocytes. The primary spermatocyte divides meiotically into two secondary spermatocytes; each secondary spermatocyte divides into two equal haploid spermatids by Meiosis II. The spermatids are transformed into spermatozoa (sperm) by the process of spermiogenesis. These develop into mature spermatozoa, also known as sperm cells. Thus, the primary spermatocyte gives rise to two cells, the secondary spermatocytes, and the two secondary spermatocytes by their subdivision produce four spermatozoa and four haploid cells.
Male infertility refers to a male's inability to cause pregnancy in a fertile female. In humans it accounts for 40–50% of infertility. It affects approximately 7% of all men. Male infertility is commonly due to deficiencies in the semen, and semen quality is used as a surrogate measure of male fecundity.
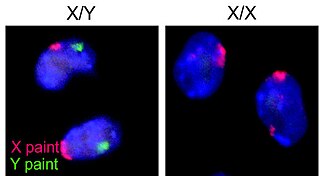
Azoospermia factor (AZF) refers to one of several proteins or their genes, which are coded from the AZF region on the human male Y chromosome. Deletions in this region are associated with inability to produce sperm. Subregions within the AZF region are AZFa, AZFb and AZFc. AZF microdeletions are one of the major causes of male infertility for azoospermia and severe oligozoospermia males. AZF is the term used by the HUGO Gene Nomenclature Committee.
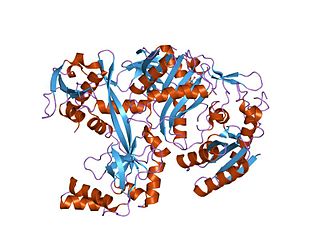
Piwi genes were identified as regulatory proteins responsible for stem cell and germ cell differentiation. Piwi is an abbreviation of P-elementInduced WImpy testis in Drosophila. Piwi proteins are highly conserved RNA-binding proteins and are present in both plants and animals. Piwi proteins belong to the Argonaute/Piwi family and have been classified as nuclear proteins. Studies on Drosophila have also indicated that Piwi proteins have slicer activity conferred by the presence of the Piwi domain. In addition, Piwi associates with heterochromatin protein 1, an epigenetic modifier, and piRNA-complementary sequences. These are indications of the role Piwi plays in epigenetic regulation. Piwi proteins are also thought to control the biogenesis of piRNA as many Piwi-like proteins contain slicer activity which would allow Piwi proteins to process precursor piRNA into mature piRNA.
CPEB, or cytoplasmic polyadenylation element binding protein, is a highly conserved RNA-binding protein that promotes the elongation of the polyadenine tail of messenger RNA. CPEB most commonly activates the target RNA for translation, but can also act as a repressor, dependent on its phosphorylation state. In animals, CPEB is expressed in several alternative splicing isoforms that are specific to particular tissues and functions, including the self-cleaving Mammalian CPEB3 ribozyme. CPEB was first identified in Xenopus oocytes and associated with meiosis; a role has also been identified in the spermatogenesis of Caenorhabditis elegans.

Polyadenylate-binding protein 1 is a protein that in humans is encoded by the PABPC1 gene. The protein PABP1 binds mRNA and facilitates a variety of functions such as transport out of the nucleus, degradation, translation, and stability. There are two separate PABP1 proteins, one which is located in the nucleus (PABPN1) and the other which is found in the cytoplasm (PABPC1). The location of PABP1 affects the role of that protein and its function with RNA.

Deleted in azoospermia-like is a protein that in humans is encoded by the DAZL gene.

Deleted in azoospermia 1, also known as DAZ1, is a protein which in humans is encoded by the DAZ1 gene.
Ubiquitin specific peptidase 9, Y-linked , also known as USP9Y, is an enzyme which in humans is encoded by the USP9Y gene. It is required for sperm production. This enzyme is a member of the peptidase C19 family and is similar to ubiquitin-specific proteases, which cleave the ubiquitin moiety from ubiquitin-fused precursors and ubiquitinylated proteins.
Deleted in azoospermia protein 3 is a protein that in humans is encoded by the DAZ3 gene.
Deleted in azoospermia protein 2 is a protein that in humans is encoded by the DAZ2 gene.

DAZ-associated protein 1 is a protein that in humans is encoded by the DAZAP1 gene.
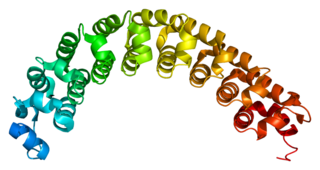
Pumilio homolog 2 is an RNA-binding protein that in humans is encoded by the PUM2 gene.

RNA-binding motif protein, Y chromosome, family 1 member A1/C is a protein that in humans is encoded by the RBMY1A1 gene.
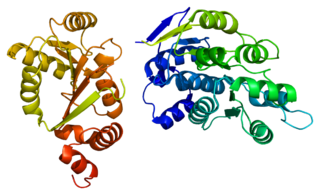
ATP-dependent RNA helicase DDX3Y is an enzyme that in humans is encoded by the DDX3Y gene.

Protein boule-like is a protein that in humans is encoded by the BOLL gene.

Sperm-associated antigen 8 is a protein that in humans is encoded by the SPAG8 gene.

DAZ-associated protein 2 is a protein that in humans is encoded by the DAZAP2 gene.
Vasa is an RNA binding protein with an ATP-dependent RNA helicase that is a member of the DEAD box family of proteins. The vasa gene, is essential for germ cell development and was first identified in Drosophila melanogaster, but has since been found to be conserved in a variety of vertebrates and invertebrates including humans. The Vasa protein is found primarily in germ cells in embryos and adults, where it is involved in germ cell determination and function, as well as in multipotent stem cells, where its exact function is unknown.
References
- 1 2 3 4 Fu XF, Cheng SF, Wang LQ, Yin S, De Felici M, Shen W (2015). "DAZ Family Proteins, Key Players for Germ Cell Development". International Journal of Biological Sciences. 11 (10): 1226–35. doi:10.7150/ijbs.11536. PMC 4551758 . PMID 26327816.
- 1 2 3 4 5 6 Reynolds N, Cooke HJ (January 2005). "Role of the DAZ genes in male fertility". Reproductive Biomedicine Online. 10 (1): 72–80. doi: 10.1016/s1472-6483(10)60806-1 . PMID 15705297.
- ↑ Dhanoa JK, Mukhopadhyay CS, Arora JS (July 2016). "Y-chromosomal genes affecting male fertility: A review". Veterinary World. 9 (7): 783–91. doi:10.14202/vetworld.2016.783-791. PMC 4983133 . PMID 27536043.
- ↑ Goss DJ, Kleiman FE (March 2013). "Poly(A) binding proteins: are they all created equal?". Wiley Interdisciplinary Reviews: RNA. 4 (2): 167–79. doi:10.1002/wrna.1151. PMC 3580857 . PMID 23424172.
- 1 2 3 4 5 Vangompel MJ, Xu EY (January 2011). "The roles of the DAZ family in spermatogenesis: More than just translation?". Spermatogenesis. 1 (1): 36–46. doi:10.4161/spmg.1.1.14659. PMC 3329454 . PMID 22523742.
- ↑ Yen PH (June 2004). "Putative biological functions of the DAZ family". International Journal of Andrology. 27 (3): 125–9. doi: 10.1111/j.1365-2605.2004.00469.x . PMID 15139965.