
The cytoskeleton is a complex, dynamic network of interlinking protein filaments present in the cytoplasm of all cells, including those of bacteria and archaea. In eukaryotes, it extends from the cell nucleus to the cell membrane and is composed of similar proteins in the various organisms. It is composed of three main components: microfilaments, intermediate filaments, and microtubules, and these are all capable of rapid growth or disassembly depending on the cell's requirements.

Microvilli are microscopic cellular membrane protrusions that increase the surface area for diffusion and minimize any increase in volume, and are involved in a wide variety of functions, including absorption, secretion, cellular adhesion, and mechanotransduction.

Actin is a family of globular multi-functional proteins that form microfilaments in the cytoskeleton, and the thin filaments in muscle fibrils. It is found in essentially all eukaryotic cells, where it may be present at a concentration of over 100 μM; its mass is roughly 42 kDa, with a diameter of 4 to 7 nm.

Tropomyosin is a two-stranded alpha-helical, coiled coil protein found in many animal and fungal cells. In animals, it is an important component of the muscular system which works in conjunction with troponin to regulate muscle contraction. It is present in smooth and striated muscle tissues, which can be found in various organs and body systems, including the heart, blood vessels, respiratory system, and digestive system. In fungi, tropomyosin is found in cell walls and helps maintain the structural integrity of cells.

In mammalian cells, vinculin is a membrane-cytoskeletal protein in focal adhesion plaques that is involved in linkage of integrin adhesion molecules to the actin cytoskeleton. Vinculin is a cytoskeletal protein associated with cell-cell and cell-matrix junctions, where it is thought to function as one of several interacting proteins involved in anchoring F-actin to the membrane.
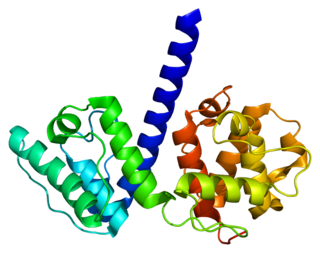
Plectin is a giant protein found in nearly all mammalian cells which acts as a link between the three main components of the cytoskeleton: actin microfilaments, microtubules and intermediate filaments. In addition, plectin links the cytoskeleton to junctions found in the plasma membrane that structurally connect different cells. By holding these different networks together, plectin plays an important role in maintaining the mechanical integrity and viscoelastic properties of tissues.

Cortactin is a monomeric protein located in the cytoplasm of cells that can be activated by external stimuli to promote polymerization and rearrangement of the actin cytoskeleton, especially the actin cortex around the cellular periphery. It is present in all cell types. When activated, it will recruit Arp2/3 complex proteins to existing actin microfilaments, facilitating and stabilizing nucleation sites for actin branching. Cortactin is important in promoting lamellipodia formation, invadopodia formation, cell migration, and endocytosis.
Actinin is a microfilament protein. The functional protein is an anti-parallel dimer, which cross-links the thin filaments in adjacent sarcomeres, and therefore coordinates contractions between sarcomeres in the horizontal axis. Alpha-actinin is a part of the spectrin superfamily. This superfamily is made of spectrin, dystrophin, and their homologous and isoforms. In non-muscle cells, it is found by the actin filaments and at the adhesion sites.The lattice like arrangement provides stability to the muscle contractile apparatus. Specifically, it helps bind actin filaments to the cell membrane. There is a binding site at each end of the rod and with bundles of actin filaments.

ROCK1 is a protein serine/threonine kinase also known as rho-associated, coiled-coil-containing protein kinase 1. Other common names are ROKβ and P160ROCK. ROCK1 is a major downstream effector of the small GTPase RhoA and is a regulator of the actomyosin cytoskeleton which promotes contractile force generation. ROCK1 plays a role in cancer and in particular cell motility, metastasis, and angiogenesis.

Tight junction protein ZO-1 also known as Zonula Occludens-1 (ZO-1), is a 220-kD peripheral membrane protein that is encoded by the TJP1 gene in humans. It belongs to the family of zonula occludens proteins, which are tight junction-associated proteins and of which, ZO-1 is the first to be cloned. It was first isolated in 1986 by Stevenson and Goodenough using a monoclonal antibody raised in rodent liver to recognise a 225-kD polypeptide in whole liver homogenates and in tight junction-enriched membrane fractions. It has a role as a scaffold protein which cross-links and anchors Tight Junction (TJ) strand proteins, which are fibril-like structures within the lipid bilayer, to the actin cytoskeleton.
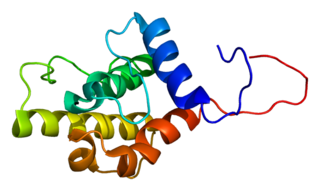
Plastin-2 is a protein that in humans is encoded by the LCP1 gene.

Spectrin beta chain, brain 1 is a protein that in humans is encoded by the SPTBN1 gene.

Myosin X, also known as MYO10, is a protein that in humans is encoded by the MYO10 gene.

Rho-associated protein kinase (ROCK) is a kinase belonging to the AGC family of serine-threonine specific protein kinases. It is involved mainly in regulating the shape and movement of cells by acting on the cytoskeleton.
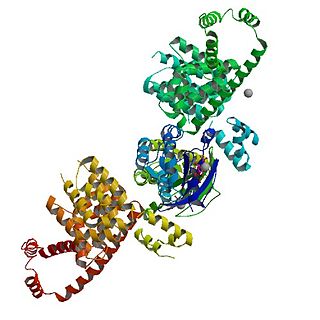
mDia1 is a member of the protein family called the formins and is a Rho effector. It is the mouse version of the diaphanous homolog 1 of Drosophila. mDia1 localizes to cells' mitotic spindle and midbody, plays a role in stress fiber and filopodia formation, phagocytosis, activation of serum response factor, formation of adherens junctions, and it can act as a transcription factor. mDia1 accelerates actin nucleation and elongation by interacting with barbed ends of actin filaments. The gene encoding mDia1 is located on Chromosome 18 of Mus musculus and named Diap1.
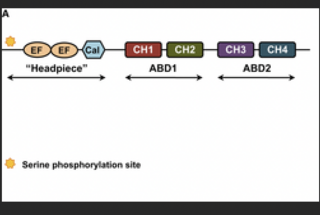
Plastin is part of a family of actin-bundling proteins, specifically the α-actinin family of actin-binding protein, which are found in many lifeforms, from humans and other animals to plants and yeasts. These proteins are known to cross-link actin filaments into bundles for various cell purposes.

In molecular biology, the IMD domain is a BAR-like domain of approximately 250 amino acids found at the N-terminus in the insulin receptor tyrosine kinase substrate p53 (IRSp53/BAIAP2) and in the evolutionarily related IRSp53/MIM (MTSS1) family. In IRSp53, a ubiquitous regulator of the actin cytoskeleton, the IMD domain acts as conserved F-actin bundling domain involved in filopodium formation. Filopodium-inducing IMD activity is regulated by Cdc42 and Rac1 and is SH3-independent. The IRSp53/MIM family is a novel F-actin bundling protein family that includes invertebrate relatives:

Calponin 2 is a protein that in humans is encoded by the CNN2 gene.

Calponin 3. acidic is a protein that in humans is encoded by the CNN3 gene.
Calponin 1 is a basic smooth muscle protein that in humans is encoded by the CNN1 gene.




