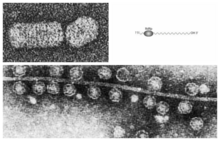
A DNA virus is a virus that has a genome made of deoxyribonucleic acid (DNA) that is replicated by a DNA polymerase. They can be divided between those that have two strands of DNA in their genome, called double-stranded DNA (dsDNA) viruses, and those that have one strand of DNA in their genome, called single-stranded DNA (ssDNA) viruses. dsDNA viruses primarily belong to two realms: Duplodnaviria and Varidnaviria, and ssDNA viruses are almost exclusively assigned to the realm Monodnaviria, which also includes some dsDNA viruses. Additionally, many DNA viruses are unassigned to higher taxa. Reverse transcribing viruses, which have a DNA genome that is replicated through an RNA intermediate by a reverse transcriptase, are classified into the kingdom Pararnavirae in the realm Riboviria.
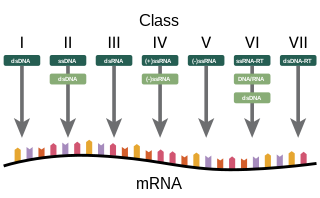
Baltimore classification is a system used to classify viruses based on their manner of messenger RNA (mRNA) synthesis. By organizing viruses based on their manner of mRNA production, it is possible to study viruses that behave similarly as a distinct group. Seven Baltimore groups are described that take into consideration whether the viral genome is made of deoxyribonucleic acid (DNA) or ribonucleic acid (RNA), whether the genome is single- or double-stranded, and whether the sense of a single-stranded RNA genome is positive or negative.

Positive-strand RNA viruses are a group of related viruses that have positive-sense, single-stranded genomes made of ribonucleic acid. The positive-sense genome can act as messenger RNA (mRNA) and can be directly translated into viral proteins by the host cell's ribosomes. Positive-strand RNA viruses encode an RNA-dependent RNA polymerase (RdRp) which is used during replication of the genome to synthesize a negative-sense antigenome that is then used as a template to create a new positive-sense viral genome.

Ortervirales is an order that contains all accepted species of single-stranded RNA viruses that replicate through a DNA intermediate and all accepted species of double-stranded DNA viruses that replicate through an RNA intermediate . The name is derived from the reverse of retro.
Yuyuevirus is a genus of negative-strand RNA viruses which infect invertebrates. Member viruses have bisegmented genomes. It is the only genus in the family Yueviridae, which in turn is the only family in the order Goujianvirales and class Yunchangviricetes. Two species are recognized: Beihai yuyuevirus and Shahe yuyuevirus.

Articulavirales is an order of segmented negative-strand RNA viruses which infect invertebrates and vertebrates. It includes the family of influenza viruses which infect humans. It is the only order of viruses in the monotypic class Insthoviricetes. The order contains two families and eight genera.

Monjiviricetes is a class of negative-strand RNA viruses which infect fungi, plants, invertebrates, and vertebrates. The name is a portmanteau of the two orders within the class, Mononegavirales and Jingchuvirales and the suffix for a virus class -viricetes.
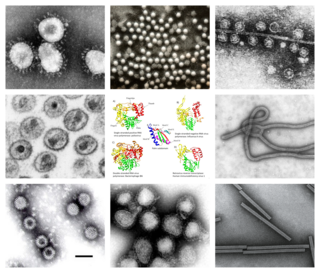
Riboviria is a realm of viruses that includes all viruses that use a homologous RNA-dependent polymerase for replication. It includes RNA viruses that encode an RNA-dependent RNA polymerase, as well as reverse-transcribing viruses that encode an RNA-dependent DNA polymerase. RNA-dependent RNA polymerase (RdRp), also called RNA replicase, produces RNA from RNA. RNA-dependent DNA polymerase (RdDp), also called reverse transcriptase (RT), produces DNA from RNA. These enzymes are essential for replicating the viral genome and transcribing viral genes into messenger RNA (mRNA) for translation of viral proteins.
In virology, realm is the highest taxonomic rank established for viruses by the International Committee on Taxonomy of Viruses (ICTV), which oversees virus taxonomy. Six virus realms are recognized and united by specific highly conserved traits:
Smacoviridae is a family of single-stranded DNA viruses. The genomes of this family are small. The name Smacoviridae stands for 'small circular genome virus'. The genomes are circular single-stranded DNA and encode rolling-circle replication initiation proteins (Rep) and unique capsid proteins. As of 2021, 12 genera and 84 species are recognized in this family. The viruses in this taxon were isolated from faecal samples from insects and vertebrates by metagenomic methods. Little is known about their biology.

Varidnaviria is a realm of viruses that includes all DNA viruses that encode major capsid proteins that contain a vertical jelly roll fold. The major capsid proteins (MCP) form into pseudohexameric subunits of the viral capsid, which stores the viral deoxyribonucleic acid (DNA), and are perpendicular, or vertical, to the surface of the capsid. Apart from this, viruses in the realm also share many other characteristics, such as minor capsid proteins (mCP) with the vertical jelly roll fold, an ATPase that packages viral DNA into the capsid, and a DNA polymerase that replicates the viral genome.

Bamfordvirae is a kingdom of viruses. This kingdom is recognized for its use of double jelly roll major capsid proteins. It was formerly known as the PRD1-adenovirus lineage. The kingdom is named after Dennis H. Bamford who first promoted the evolutionary unity of all viruses encoding double jelly-roll major capsid proteins.

Orthornavirae is a kingdom of viruses that have genomes made of ribonucleic acid (RNA), those genomes encoding an RNA-dependent RNA polymerase (RdRp). The RdRp is used to transcribe the viral RNA genome into messenger RNA (mRNA) and to replicate the genome. Viruses in this kingdom also share a number of characteristics involving evolution, including high rates of genetic mutations, recombinations, and reassortments.
Duplornaviricota is a phylum of RNA viruses, which contains all double-stranded RNA viruses, except for those in phylum Pisuviricota. Characteristic of the group is a viral capsid composed of 60 homo- or heterodimers of capsid protein on a pseudo-T=2 lattice. Duplornaviruses infect both prokaryotes and eukaryotes. The name of the group derives from Italian duplo which means double, rna for the type of virus, and -viricota which is the suffix for a virus phylum.
Kitrinoviricota is a phylum of RNA viruses that includes all positive-strand RNA viruses that infect eukaryotes and are not members of the phylum Pisuviricota or Lenarviricota. The name of the group derives from Greek κίτρινος (kítrinos), which means yellow, and -viricota, which is the suffix for a virus phylum.

Pisuviricota is a phylum of RNA viruses that includes all positive-strand and double-stranded RNA viruses that infect eukaryotes and are not members of the phylum Kitrinoviricota,Lenarviricota or Duplornaviricota. The name of the group is a syllabic abbreviation of “picornavirus supergroup” with the suffix -viricota, indicating a virus phylum. Phylogenetic analyses suggest that Birnaviridae and Permutotetraviridae, both currently unassigned to a phylum in Orthornavirae, also belong to this phylum and that both are sister groups. Another proposed family of the phylum is unassigned Polymycoviridae in Riboviria.

Stelpaviricetes is a class of non-enveloped, positive-strand RNA viruses which infect plants and animals. Characteristic of the group is a VPg protein attached to the 5'-end of the genome and a conserved 3C-like protease from the PA clan of proteases for processing the translated polyprotein. The name of the group is a syllabic abbreviation of member orders "stellavirales, patatavirales" with the suffix -viricetes denoting a virus class.

An archaeal virus is a virus that infects and replicates in archaea, a domain of unicellular, prokaryotic organisms. Archaeal viruses, like their hosts, are found worldwide, including in extreme environments inhospitable to most life such as acidic hot springs, highly saline bodies of water, and at the bottom of the ocean. They have been also found in the human body. The first known archaeal virus was described in 1974 and since then, a large diversity of archaeal viruses have been discovered, many possessing unique characteristics not found in other viruses. Little is known about their biological processes, such as how they replicate, but they are believed to have many independent origins, some of which likely predate the last archaeal common ancestor (LACA).

Portogloboviridae is a family of DNA viruses that infect archaea. It is a proposed family of the realm Varidnaviria. Viruses in the family are related to Halopanivirales. The capsid proteins of these viruses and their characteristics are of evolutionary importance for the origin of the other Varidnaviria viruses since they seem to retain primordial characters.

Adnaviria is a realm of viruses that includes archaeal viruses that have a filamentous virion and a linear, double-stranded DNA genome. The genome exists in A-form (A-DNA) and encodes a dimeric major capsid protein (MCP) that contains the SIRV2 fold, a type of alpha-helix bundle containing four helices. The virion consists of the genome encased in capsid proteins to form a helical nucleoprotein complex. For some viruses, this helix is surrounded by a lipid membrane called an envelope. Some contain an additional protein layer between the nucleoprotein helix and the envelope. Complete virions are long and thin and may be flexible or a stiff like a rod.