
The haptophytes, classified either as the Haptophyta, Haptophytina or Prymnesiophyta, are a clade of algae.

Euglena is a genus of single cell flagellate eukaryotes. It is the best known and most widely studied member of the class Euglenoidea, a diverse group containing some 54 genera and at least 200 species. Species of Euglena are found in fresh water and salt water. They are often abundant in quiet inland waters where they may bloom in numbers sufficient to color the surface of ponds and ditches green (E. viridis) or red (E. sanguinea).
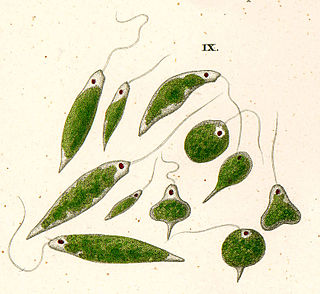
Euglenids are one of the best-known groups of flagellates, which are excavate eukaryotes of the phylum Euglenophyta and their cell structure is typical of that group. They are commonly found in freshwater, especially when it is rich in organic materials, with a few marine and endosymbiotic members. Many euglenids feed by phagocytosis, or strictly by diffusion. A monophyletic group consisting of the mixotrophic Rapaza viridis and the two groups Eutreptiales and Euglenales have chloroplasts and produce their own food through photosynthesis. This group is known to contain the carbohydrate paramylon.

Chlamydomonas is a genus of green algae consisting of about 150 species of unicellular flagellates, found in stagnant water and on damp soil, in freshwater, seawater, and even in snow as "snow algae". Chlamydomonas is used as a model organism for molecular biology, especially studies of flagellar motility and chloroplast dynamics, biogenesis, and genetics. One of the many striking features of Chlamydomonas is that it contains ion channels (channelrhodopsins) that are directly activated by light. Some regulatory systems of Chlamydomonas are more complex than their homologs in Gymnosperms, with evolutionarily related regulatory proteins being larger and containing additional domains.

Chromista is a proposed but polyphyletic biological kingdom consisting of single-celled and multicellular eukaryotic species that share similar features in their photosynthetic organelles (plastids). It includes all eukaryotes whose plastids contain chlorophyll c and are surrounded by four membranes. If the ancestor already possessed chloroplasts derived by endosymbiosis from red algae, all non-photosynthetic Chromista have secondarily lost the ability to photosynthesise. Its members might have arisen independently as separate evolutionary groups from the last eukaryotic common ancestor.
Cryptomonas is the name-giving genus of the Cryptomonads established by German biologist Christian Gottfried Ehrenberg in 1831. The algae are common in freshwater habitats and brackish water worldwide and often form blooms in greater depths of lakes. The cells are usually brownish or greenish in color and are characteristic of having a slit-like furrow at the anterior. They are not known to produce any toxins. They are used to feed small zooplankton, which is the food source for small fish in fish farms. Many species of Cryptomonas can only be identified by DNA sequencing. Cryptomonas can be found in several marine ecosystems in Australia and South Korea.

Tetraselmis is a genus of phytoplankton. Tetraselmis is a green algal genus within the order Chlorodendrales, and they are characterized by their intensely-colored green chloroplast, their flagellated cell bodies, the presence of a pyrenoid within the chloroplast, and a scale-produced thecal-wall. Species within this genus are found in both marine and freshwater ecosystems across the globe; their habitat range is mainly limited by water depth due to their photosynthetic nature. Thus, they live in diverse water environments if enough nutrients and light are available for net photosynthetic activity. Tetraselmis species have proven to be useful for both research and industry. Tetraselmis species have been studied for understanding plankton growth rates, and recently a colonial species is being used to gain an understanding of multicellularity evolution. Additionally, many species are currently being examined for their use as biofuels due to their high lipid content.
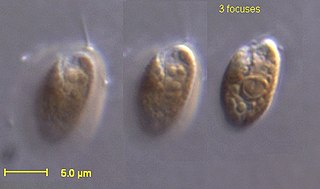
The cryptophyceae are a class of algae, most of which have plastids. About 220 species are known, and they are common in freshwater, and also occur in marine and brackish habitats. Each cell is around 10–50 μm in size and flattened in shape, with an anterior groove or pocket. At the edge of the pocket there are typically two slightly unequal flagella.

Bodo is a genus of microscopic kinetoplastids, flagellate excavates first described in 1831 by Christian Gottfried Ehrenberg. The genus is small, as it has recently been redefined to include only four species. Bodo includes free-living, phagotrophic organisms that can be found in many marine and freshwater environments as well as some terrestrial environments. Being phagotrophic, Bodo feeds on bacteria and other microorganisms that it finds while swimming through its water-based habitats. The swimming-like movement is facilitated by the two unequal flagella that Bodo possesses which arise from an anteriorly located flagellar pocket. Bodo is roughly bean-shaped and is often missed in samples from water or terrestrial environments due to its small size.

Eustigmatophytes are a small group of eukaryotic forms of algae that includes marine, freshwater and soil-living species.
Pavlovaceae is a family of haptophytes. It is the only family in the order Pavlovales, which is the only order in the class Pavlovophyceae. It contains four genera, Diacronema, Exanthemachrysis, Pavlova and Rebecca.
Rhodomonas is a genus of cryptomonads. It is characterized by its red colour, the square-shaped plates of its inner periplast, its short furrow ending in a gullet, and a distinctly shaped chloroplast closely associated with its nucleomorph. Historically, Rhodomonas was characterized by its red chloroplast alone, but this no longer occurs as its taxonomy has become increasingly based on molecular and cellular data. Currently, there is some debate about the taxonomic validity of Rhodomonas as a genus and further research is needed to verify its taxonomic status. Rhodomonas is typically found in marine environments, although freshwater reports exist. It is commonly used as a live feed for various aquaculture species.
Diacronema is a genus of haptophytes.

Chrysochromulina is a genus of haptophytes. This phytoplankton is distributed globally in brackish and marine waters across approximately 60 known species. All Chrysochromulina species are phototrophic, however some have been shown to be mixotrophic, including exhibiting phagotrophy under certain environmental conditions. The cells are small, characterized by having scales, and typically observed using electron microscopy. Some species, under certain environmental conditions have been shown to produce toxic compounds that are harmful to larger marine life including fish.

Thaumatomastix is a protist genus of the order Thaumatomonadida, within the phylum Cercozoa and the class Imbricatea. Its species are aquatic, feeding on algae and appearing in waters of a wide range of temperatures and salinities, and are 15-50 micrometers long. They can interchange between flagellated and amoeboid forms, and are notable for being covered in both spiny and flattened siliceous scales.

Phacus is a genus of unicellular excavates, of the phylum Euglenozoa, characterized by its flat, leaf-shaped structure, and rigid cytoskeleton known as a pellicle. These eukaryotes are mostly green in colour, and have a single flagellum that extends the length of their body. They are morphologically very flat, rigid, leaf-shaped, and contain many small discoid chloroplasts.
Haptista is a proposed group of protists made up of centrohelids and haptophytes. Phylogenomic studies indicate that Haptista, together with Ancoracysta twista, forms a sister clade to the SAR+Telonemia supergroup, but it may also be sister to the Cryptista (+Archaeplastida). It is thus one of the earliest diverging Diaphoretickes.
Torodinium (ˌtɔɹoʊˈdɪniəm) is a genus of unarmored dinoflagellates and comprises two species, Torodinium robustum and the type species Torodinium teredo. The establishment of Torodinium, as well as the characterization of the majority of its morphology, occurred in 1921 and further advances since have been slow. Lack of research is largely due to its extremely fragile and easily deformed nature, which also renders fossil records implausible. The genus was originally characterized by torsion of the sulcus and a posterior cingulum. Since then, new distinctive features have been discovered including an extremely reduced hyposome, a longitudinally ribbed episome, and a canal on the dextro-lateral side. Further investigation into the function of many anatomical features is still necessary for this genus.
Rhodelphis is a single-celled archaeplastid that lives in aquatic environments and is the sister group to red algae and possibly Picozoa. While red algae have no flagellated stages and are generally photoautotrophic, Rhodelphis is a flagellated predator containing a non-photosynthetic plastid. This group is important to the understanding of plastid evolution because they provide insight into the morphology and biochemistry of early archaeplastids. Rhodelphis contains a remnant plastid that is not capable of photosynthesis, but may play a role in biochemical pathways in the cell like heme synthesis and iron-sulfur clustering. The plastid does not have a genome, but genes are targeted to it from the nucleus. Rhodelphis is ovoid with a tapered anterior end bearing two perpendicularly-oriented flagella.
Cryptoglena(/ˌkɹɪptoʊˈgliːnə/) is a genus of photosynthetic euglenids that was first described in 1831 by Christian Gottfried Ehrenberg. Today, its circumscription is controversial: Bicudo and Menezes consider twenty-one species as Cryptoglena, of which, nine are uncertain. Cryptoglena species are water-based, living in both freshwater and marine environments. They are biflagellated, with one internal flagellum and one external flagellum, which allows movement through environments as demonstrated by Kim and Shin in the species C. pigra. The cells of Cryptoglena resemble a coffee bean, as they have a groove that runs the length of the cell on one side and makes them U-shaped in cross section. They are ovoid in shape and are small, with the larger cells being on average 25 x 15 μm. After being first described in 1831, little work was done on the genus until the late 1970s and early 1980s, after the scanning electron microscope completed development and was implemented into laboratories. Work then proceeded with the developments of molecular biology, which allows for classifications based on DNA sequences. For Cryptoglena the main DNA used for classification are small subunit (SSU) and large subunit (LSU) rDNA.