Arctodus is an extinct genus of short-faced bear that inhabited North America during the Pleistocene (~2.5 Mya until 12,800 years ago). There are two recognized species: the lesser short-faced bear (Arctodus pristinus) and the giant short-faced bear (Arctodus simus). Of these species, A. simus was larger, is known from more complete remains, and is considered one of the most charismatic of North America's megafauna. A. pristinus was largely restricted to the Early Pleistocene of the Eastern United States, whereas A. simus had a broader range, with most finds being from the Late Pleistocene of the United States, Mexico and Canada. A. simus evolved from A. pristinus, but both species likely overlapped in the Middle Pleistocene. Both species are relatively rare in the fossil record.
Today considered to be an enormous omnivore, Arctodus simus is believed to be one of the largest known terrestrial carnivorans that has ever existed. However, Arctodus, like other bears, was highly sexually dimorphic- adult A. simus ranged between 300 and 950 kilograms (660 and 2,090lb), with females clustering at ≤500 kilograms (1,100lb), and males around 800 kilograms (1,800lb). The largest males stood at 1.67 metres (5ft 6in) at the shoulder, and up to 3.4 metres (11ft) tall on their rear legs. Studies suggest that Arctodus simus browsed on C3 vegetation and consumed browsing herbivores such as deer, camelids, and tapir. A. simus preferred temperate open woodlands but was an adaptable species, taking advantage of many habitats and feeding opportunities.
Arctodus belongs to the Tremarctinae subfamily of bears, which are endemic to the Americas. Of these short-faced bears, Arctodus was the most widespread in North America. However, both species went extinct in the Pleistocene. A. pristinus went extinct around 300,000 years ago, with A. simus disappearing ~12,800 years ago in the Late Pleistocene extinctions. The cause behind these extinctions is unclear, but in the case of A. pristinus, this was likely due to climate change and competition with other ursids, such as the black bear and Tremarctos floridanus. A. simus likely went extinct due to ecological collapse disrupting the vegetation and prey it relied on.
Arctodus was first described by Joseph Leidy in 1854, with finds of A. pristinus from the Ashley Phosphate Beds, South Carolina.[1][2][3] The scientific name of the genus, Arctodus, derives from Greek, and means "bear tooth". The first fossils of Arctodus simus were found in the Potter Creek Cave, Shasta County, California, by J. A. Richardson in 1878, and were initially described as Arctotherium simum by Edward Drinker Cope in 1879.[4][5][6] Historically, all specimens were grouped together under A. pristinus, until a revision by Björn Kurtén in 1967.[7]
In the 19th and early 20th centuries, specimens of Arctodus were occasionally referred to Arctotherium, and vice versa.[6][8][9][10][11] However, today neither genera are considered to have overlapped, with the closest point of contact being México, with the giant Arctodus simus in Valsequillo, Puebla,[7][12][13] and the smaller Arctotherium wingei in the Yucatán Peninsula.[14] Other early researchers believed Arctodus to be a sister lineage of the agriotheriinIndarctos.[15] Sometimes described as the "American cave bear",[4]Arctodus should not be mistaken for the similarly large Eurasian cave bear (Ursus spelaeus). As an ursine, the Eurasian cave bear last shared a common ancestor with the tremarctine Arctodus circa 13.4 million years ago.[16]
Fossils of Arctodus pristinus can be confused with the similarly sized, partially contemporaneous short-faced bear, Tremarctos floridanus.[1]Arctodus has higher crowned and considerably larger teeth than its relative Tremarctos. A. pristinus can be distinguished by broader and taller molars on average, but as they are often worn, differentiation can be difficult.[2] Moreover, diagnosing isolated A. simus remains (such as femora, scapulae, certain vertebrae, ribs, podials) from brown bears can be challenging, as some large brown bears overlap in dimensions with small Arctodus simus.[7] Beyond standard differences between tremarctine and ursine bears, A. simus has a more anterior protocone & extended enamel ridge forming a shearing blade on the maxillary P4. The molars are also shorter & broader in Arctodus than brown bears.[17]
Arctodus belongs to the subfamily Tremarctinae, which appeared in North America during the earliest parts of the late Miocene epoch in the form of Plionarctos. The medium-sized Arctodus pristinus,Tremarctos floridanus and Arctotherium sp. evolved from Plionarctos in the Blancan age of North America.[2][18][19] The genetic divergence date for Arctodus is ~5.5 million years ago,[16] around the Miocene-Pliocene boundary (~5 Mya), when tremarctine bears, along with other ursids, experienced an explosive radiation in diversity, as C4 vegetation (grasses) and open habitats dominated. The world experienced a major temperature drop and increased seasonality, and a faunal turnover which extinguished 70–80% of North American genera.[20][21]
Arctodus pristinus was mostly restricted to the more densely forested thermal enclave in eastern North America.[22][23]A. pristinus has the greatest concentration of fossils in Florida,[2] with the earliest finds being from the Late Blancan Kissimmee River6 (2.7 - 2.2 Mya) and Santa Fe River1 sites.[1][2][24] During the early Irvingtonian age (~1.36 million years ago), a western population of A. pristinus evolved into the enormous A. simus, being first recorded from the Irvington type locality in California.[7][25] Correspondingly, A. simus is most plentiful from western North America,[26][27] albeit preferring mixed habitat such as temperate open woodlands.[28][29][30][31][32] Their ranges may have met in the Middle Pleistocene of Kansas,[7] with A. simus migrating east in the Late Pleistocene (around the extinction of A. pristinus).[19][33] Although both Arctodus species co-inhabited North America for at least a million years during the Middle Pleistocene (A. pristinus went extinct ~300,000 BP), there is no direct evidence of overlap or competition in the fossil record as of yet, as both species established largely separate ranges.[19]
Irvingtonian age (1,900,000 BP - 250,000 BP) specimens of Arctodus simus are particularly sparse. Finds are mostly from California, with additional remains from Texas, Kansas, Nebraska, and Montana.[34][35][36] However, A. simus became a pan-continental species in the Rancholabrean (Late Pleistocene), sharing that distinction with the black bear.[25][31] Despite Arctodus simus' large temporal and geographic range, fossil remains are comparatively rare (109 finds as of 2010, in otherwise well-sampled localities).[19][16]
Description
Size
Restoration of Arctodus simus.
Arctodus pristinus
Around the size of grizzly bears, A. pristinus specimens closely overlap the size of Tremarctos floridanus, with some males of A. pristinus overlapping in size with the females of A. simus.[1]A. pristinus individuals recovered from the Leisey Shell Pit, Florida, were calculated to an average of ~133 kilograms (293lb).[37] However, the dimensions of some individuals from Port Kennedy Bone Cave and Aguascalientes suggest that northern and western A. pristinus may have been larger than Floridan A. pristinus,[7] being up to 400 kilograms (880lb).[38]
Arctodus simus
Some A. simus individuals might have been the largest land-dwelling specimens of Carnivora that ever lived in North America. Standing up on its hind legs, A. simus stood 2.4–3.4m (8–11ft),[39][40] with a maximum vertical arm reach of 4.3 metres (14ft).[41] When walking on all fours, A. simus stood 1–1.67m (3.3–5.5ft) high at the shoulder, with the largest males being tall enough to look an adult human in the eye.[42][40][43] The average weight of A. simus was ~625 kilograms (1,378lb), with the maximum recorded at 957 kilograms (2,110lb).[44][37]
Studies
In a 2010 study, the mass of six A. simus specimens was estimated; one-third of them weighed about 850kg (1,874lb), the largest from Salt Lake Valley, Utah coming in at 957kg (2,110lb), suggesting larger specimens were probably more common than previously thought. However, half the specimens were calculated to be less than 500kg (1,100lb). The weight range calculated from all examined specimens was between 957kg and 317kg (699lb).[44] Hypothetically, the largest individuals of A. simus may have approached 1,000 kilograms (2,200lb),[45] or even 1,200kg (2,600lb).[38] However, a 2006 study argued that the maximum size of Arctodus was ~555 kilograms (1,224lb), based on the largest known skull.[46] Additionally, a 1998 study calculated the average weight of Arctodus specimens from the La Brea Tar Pits at ~372 kilograms (820lb), smaller than recovered brown bear remains (~455 kilograms (1,003lb), although these remains postdate Arctodus).[47][48][49] A 1999 study by Per Christiansen calculated a mean weight of ~770 kilograms (1,700lb) from six large male A. simus specimens.[45]
There is much variation in adult size among specimens- the lack of finds, sexual dimorphism, individual variation and potentially ecomorphs could be augmenting the average size of both species of Arctodus.[2][7][50] Size differences between specimens of Arctodus simus (such as skull and long bone dimensions) led Kurtén to suggest a larger northern/central subspecies (A. s. yukonensis) and a southern subspecies (A. s. simus).[44][7][36] evolving in the Irvingtonian and Rancholabrean respectively.[36][7] However, the discovery of a very large southern Arctodus simus in Florida and New Mexico (deep within the supposed range of A. s. simus),[19][51] & possibly Rancho La Brea,[7] and notably small specimens from the Yukon and Vancouver Island,[16][17] put doubt on this designation.[52] Perceived ecomorphologies are possibly due to the low number of specimens, and sex-biased sampling.[19]
For example, only one baculum (penis bone) has been recovered from over 100 giant short-faced bear sites in North America, although it may belong to a black bear (Potter Cave).[53] None of the specimens assigned to the larger morph (A. s. yukonensis) is from a cave passage, being usually isolated remains from open sites. Furthermore, over 70% of the smaller specimens (once assigned as the A. s. simus subspecies) are from cave deposits where bacula would likely be found if present, suggesting that mostly female individuals of A. simus were using caves.[54][55][7] Therefore, in conjunction with ursid sexual dimorphism (e.g. male spectacled bears are 30% - 40% larger than females), the larger, massive Arctodus individuals are often considered male, particularly older males, with the smaller, more lightly built individuals being females.[19][56][34] Sexual dimorphism may also explain A. simus teeth (from multiple individuals at the same site) generally clustering into two sizes.[7]
Anatomy
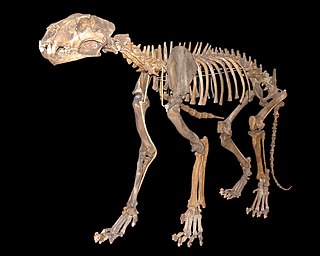
The two species of Arctodus are differentiated not only by size, but also by the shorter snout, greater prognathism, more robust teeth and longer limbs of A. simus, and the relative proportions of each species' molars and premolars. Arctodus pristinus is distinguished from A. simus smaller, narrower, and less crowded teeth. However, the morphologies of both species are otherwise very similar. As a result, differentiating Arctodus simus from Arctodus pristinus can be difficult, as male individuals of Arctodus pristinus can overlap in size with female individuals of Arctodus simus.[19][54][57]Arctodus simus superficially resembled living hyaenids in skull shape and relative lengths of the trunk, back and limbs.[46] The most nearly complete skeleton of A. simus found in the United States was unearthed in Fulton County, Indiana; the original bones are in the Field Museum of Natural History, Chicago.[42][53]
Members of the Tremarctinae subfamily of bears appear to have a disproportionately short snout compared with most modern bears, giving them the name "short-faced". Arctodus has also been argued to exhibit a wide and shortened rostrum, potentially giving Arctodus a more felid-like appearance.[36][58] Matheus suggested that a broad snout could have housed a highly developed olfactory apparatus, or accommodated a larger throat passage to bolt down large food items, akin to spotted hyenas.[59] However, this apparent shortness is an illusion caused by the deep snouts and short nasal bones of tremarctine bears compared with ursine bears; Arctodus has a deeper but not a shorter face than most living bears. This characteristic is also shared by the only living tremarctine bear, the omni-herbivorous spectacled bear.[26][44][60] Snout deepness could be variable, as specimens from Huntington Reservoir in Utah, and the Hill-Shuler locality, Texas, were noted as being distinctly "short-faced" in comparison with other Arctodus simus individuals.[61][62]
The orbits of Arctodus are proportionally small compared to the size of the skull, and somewhat laterally orientated (a characteristic of tremarctine bears), more so than actively predatory carnivorans or even the brown bear, suggesting that stereoscopic vision was not a priority.[46][60][63] The optic canal and other sphenoidal openings crowd together more in A. simus than in Ursus.[17] As with Tremarctos ornatus, specimens with a large sagittal crest were likely male, whereas females had a reduced or no sagittal crest.[2] Although there are limited samples, the middle earbones of A. simus are proportionally larger than modern ursine bears, suggesting the species was particularly attuned to low-frequency sounds.[64]
Morphologically, Arctodus simus exhibits masticular characteristics common to herbivorous bears. This includes cheek teeth with large, blunt surface areas, a deep mandible, and large mandibular muscle attachments (which are rare in carnivorous mammals). As herbivorous carnivorans such as Arctodus lack the gut microbiota to efficiently break down plant matter, these features created a high mechanical advantage of the jaw to break down plant matter via extensive chewing or grinding.[26][21][65] Although the low mandibular condyle relative to the tooth row (and therefore potential wide gape) of Arctodus simus has been inferred as an adaptation for carnivory,[36][66][17] it is also present in the omni-herbivorous spectacled bear.[44] However, both Arctodus pristinus and Tremarctos floridanus have condyles raised well above the plane of the teeth.[67] The purpose of the highly vaulted calvarium and straight cheek bones of Arctodus simus have been similarly disputed.[44]
A 2009 analysis of the mandibular morphology of tremarctine bears found notable differences between Arctodus pristinus and Arctodus simus, with A. simus specimens possessing a concave jaw, large masseter and temporalis muscles, deeper horizontal ramus and a reduced slicing dentition length when compared to A. pristinus. Instead, Arctodus simus was most similar to Arctotherium angustidens- however, both species of Arctodus and Arctotherium angustidens were still comfortably in the "omnivorous" bear cranio-morphotype.[60]
Dentition
Right lower second molar (M2) of Arctodus simus, from Rancho La Brea, California.
The premolars and first molars of Arctodus pristinus are relatively smaller and more widely spaced than those of Arctodus simus.[1][19] In A. pristinus, the features of the dentition can be quite variable, particularly the M2 molar.[2] An analysis of the Hunter-Schreger bands from the teeth of A. pristinus and A. simus demonstrated an evolutionary trend towards partially reinforced tooth enamel. This has been convergently evolved with giant pandas, agriotheriin bears, and Hemicyon.[68] The dentition of A. simus has been used as evidence of a predatory lifestyle- in particular the large canines, the high-crowned lower first molar, and the possible carnassial shear with the upper fourth premolar. However, the wearing of the molars to a relatively flat & blunt loph (suitable as a crushing platform as per modern omnivorous bears), small shear facet, and the flattened cusps across age ranges (unlike carnivores, which instead have carnassial shears) disagrees with this hypothesis.[26][69][17]
Dentition can be a poor indicator of size in A. simus, as some medium-size individuals have teeth that surpass the size of those with the largest skeletons.[7] Additionally, while A. simus evolved from the smaller A. pristinus, their teeth remained generally the same size.[7] A specimen of A. simus from the Seale Pit of the Hill-Shuler locality, Texas, with only two premolars, crowding of the anterior premolar out of line, and a wider and shorter muzzle, was suggested to be an undescribed form of Arctodus.[62]
Post-cranial
Limbs
A. simus compared with a human.
Researchers have differing interpretations on the limb morphology of Arctodus. A comprehensive 2010 study concluded that the legs of Arctodus weren't proportionally longer than modern bears would be expected to have, and that bears in general are long-limbed animals, obscured in life by their girth and fur. The study concluded the supposed "long-legged" appearance of the bear is largely an illusion created by the animal's relatively shorter back and torso. In fact, Arctodus probably had an even shorter back than other bears, due the necessary ratio between body length and body mass of the huge bear.[44][70] However, other researchers argue that the limb bones of Arctodus simus are proportionally longer than those of other bears, leading to a "gracile" appearance. Although longer, the proportions still overlap with Ursus, and the limb bones are stouter than in the large-bodied felids (Panthera). Rather than for running, these elongated limb bones may have evolved for increased locomotor efficiency during prolonged travel.[71][72] This stiff-legged, swinging gait could have been similar to a polar bear's.[73] Some researchers suggest that proportionally longer limbs may be an adaptation for increased vision over tall ground cover in an open habitat, or were used in tearing and pulling down vegetation.[26][43]
Researchers also disagree when interpreting the humerus of Arctodus simus.[71] Sorkin argued that the pronation of the forearm and the flexion of the wrist and digits, and more lightly muscled forelimbs, all of which are crucial to grasping a large prey animal with the forepaws, were probably less powerful in Arctodus than in either the brown bear or in Panthera. This is due to a weak medial epicondyle and reduced development of the pronator teres muscle.[46] The forelimb of Arctodus could have been in the early stages of cursorial evolution, being capable of more efficient and high-speed straight-line locomotion (relative to extant bears), and was possibly more adept at pursuing large prey than polar and brown bears.[74] On the other hand, some researchers argue that the epicondyles were still well developed, with this wide range of ulna rotation suggests that forearms of Arctodus were powerful and could subdue large prey.[71] The shape of the elbow joint, along with an well-developed medial epicondyle which forms an angle with the condyle, and shallower olecranon fossa, would have given Arctodus a higher degree of forelimb dexterity. Originally evolved to facilitate arboreality, other researchers believe that the terrestrial Arctodus (along with Arctotherium and the giant panda) retained this characteristic to assist in foraging for vegetation.[75][26][76]
Paws
The paws (metapodials and phalanges) of Arctodus were characteristically long, slender, and more elongated along the third and fourth digits compared to ursine bears. Arctodus' paws were therefore more symmetrical than ursine bears, whose feet have axes aligned with the most lateral (fifth) digit. Also, the first digit of Arctodus was positioned more closely and parallel to the other four digits (i.e. with straight toes, Arctodus had less lateral splaying).[70][43]
For comparison, the manus of the spectacled bear has five digits arrayed in a shallow arc (Chaquil Cave, Peru).[77][78]
However this is potentially contradicted by possible Arctodus simus trackways from near Lakeview, Oregon, with strong toe splaying, three centrally aligned & evenly spaced toes at the front, and two almost perpendicular lateral toes (80° from the axis of the foot on either side). The trackways suggest that Arctodus had an oval-shaped, undivided pad on its sole, front paws that were slightly larger than its back paws, possessed long claws, and had its hind foot overstep the forefoot when walking, like modern bears.[79] An additional A. simus paw print measuring 15cm (5.9in) long and 19cm (7.5in) wide has been recovered from White Sands National Park, New Mexico.[80][81] Some claw marks attributed to Arctodus simus at Riverbluff Cave (as they were four meters above the floor of the cave) were nearly 20cm in width.[82][83]
The presence of a partial false thumb in Arctodus simus is a characteristic shared with Tremarctos floridanus and the spectacled bear, and is possibly an ancestral trait. Absent in ursine bears, the false thumb of the spectacled bear has been suggested to assist in herbivorous food manipulation (such as bromeliads, leaves, berries, tree bark & fruits, cactus fruits & pulp, palm hearts & fronds), or arboreality.[84][21][26]
Paleopathology
Beyond carbohydrate-associated dental pathologies present in the genus,[54][85][17] extensive pathologies have been preserved on the most nearly complete skeleton of Arctodus. The leading hypothesis suggests the Fulton CountyArctodus specimen suffered from a syphilis-like (treponemal) disease, or yaws, based on the various lesions present.[42][86][87] The same individual records a pathological growth distorting the right humerus,[46] with abscesses are noted between the molars and on both ulna. Hypotheses include syphilis, osteoarthritis, a fungal infection in addition to long term syphilis, or an infected wound.[42][88] Several specimens from Fairbanks, Alaska, also exhibit either pathological growths or periodontal disease,[7] along with a healed toe bone from Big Bear Cave, Missouri.[53]
Paleobiology
Locomotion
Paul Matheus proposed that Arctodus simus may have moved in a highly efficient, moderate-speed pacing gait, more specialized than modern bears. His research concluded that the large body size, taller front legs, high shoulders, short and sloping back, and long legs of Arctodus also compounded locomotive efficiency, as these traits swelled the amount of usable elastic strain energy in the tendons, and increased stride length, making Arctodus built more for endurance than for great speed.[39][72] His calculations suggested that Arctodus likely had a top speed of 40–45 kilometres per hour (25–28mph), and based on hyaenid proportions, would shift from singlefoot locomotion to a pace at 8.5km/h (5.3mph), and would begin to gallop at 18.5km/h (11.5mph), a fairly high speed. Based on other mammals, the optimal pace speed of Arctodus would have been 13.7km/h (8.5mph). For comparison, hyenas cross country ~10km/h (6.2mph).[72] This mobility would have facilitated travelling across a large home range, which Mattson suggests may have topped 1,000 square miles (2,600km2).[53] Swimming has also been presented as a hypothesis for the colonization of Vancouver Island by Arctodus simus.[89][53]
Maturity
Examinations on a mostly full sized young individual of Arctodus simus from an Ozark cave suggest that Arctodus, like other ursids, reached sexual maturity well before full maturity. Comparisons with black bears suggest the Arctodus specimen was either 4–6 years of age if female, or 6–8 years if the specimen was male. Additionally, wear patterns on the individual's teeth are similar to a 4-6 year old Ursus americanus. Fused sutures, epiphyses, and epiphyseal plates, along with tooth eruption, have been used to determine adulthood in Arctodus.[54][16]
Legend: Main mitochondrial haplogroup of A. simus (haplotypes A, B, C, D, E, F, G)
Chiquihuite Cave A.simus
Unassigned A.simus
An examination of mitochondrial DNA sequenced from specimens of Arctodus simus from Alaska, Alberta, Ohio and the Yukon suggest an extremely low level of genetic diversity among the 23 individuals studied (≤ 44,000 14C BP), with only seven haplotypes recovered. Genetic diversity was comparable to modern endangered fauna, such as the brown kiwi and African cheetah. Explanations include a genetic bottleneck before 44,000 14C BP, or a low level of genetic diversity being a feature of a species which was primarily solitary, with a large home range and relatively small population size.[90] However, this does not entirely preclude genetic diversity in Arctodus simus, with genetic samples from Chiquihuite Cave, Zacatecas indicating a deep divergence with previously studied specimens of A. simus.[16] Additional specimens from the California Channel Islands and Wyoming have been sequenced, but are unassigned.[91]
Haplotype cladogram
Below is a cladogram exploring the relationships between the mitochondrial haplogroups of Arctodus simus. Other than the specimen from Chiquihuite Cave, all haplotypes form a single clade.[16][90]
Arctodus faeces found at South Dakota and the Yukon contain Juniperus seeds. Seed cones and berries are still important food sources for northern bears today.
The fact that Arctodus did not significantly differ in dentition or build from modern bears has led most authors to support the hypothesis that the A. simus was omnivorous, like most modern bears, and would have eaten significant amounts of plant matter.[69][92] Morphologically, Arctodus simus exhibits masticular and dental characteristics which confirms that short-faced bears such as the spectacled bear and Arctodus were adapted to and actively consumed vegetation.[26][60][21][69][65] This is affirmed by a lack of dental damage associated with carnivory amongst specimens of Arctodus.[93] Dental pathologies which have been found, such as incisor wear & supragingival dental calculus in a young individual from Missouri,[54] and cavities associated with carbohydrate consumption in individuals from the La Brea Tar Pits & Pellucidar Cave (Vancouver Island), further suggest an omnivorous diet for Arctodus simus.[85][93][17] Additional morphological adaptations include dexterous forelimbs and a partial false thumb, which would have assisted in foraging for vegetation,[76][84] along with the body size of large Arctodus simus (~1000kg) matching or exceeding the expected upper limitations for a terrestrial carnivore (based on the more restrictive energy base for a carnivorous diet).[26][94][95]
While features of Arctodus simus morphology suggest herbivory, their close phylogenetic relationship to the omni-herbivorous spectacled bear presents the possibility that these traits may be an ancestral condition of the group. A browsing diet foraged from the canopies of trees and shrubs could have been difficult with the large and flattened rostrum and incisor build of Arctodus, with an absence of digging adaptations in Arctodus' forelimbs and claws limiting rooting.[43][46] Regardless, gross tooth wear suggests consumption of plant matter in the diet of Arctodus simus.[21][26] The diet of individuals from La Brea was most similar to the spectacled bear, which consumes tough leafy matter, seeded & pitted fruits and occasional protein. Arctodus' tooth wear remained consistent throughout the Pleistocene in La Brea. This indicated a less generalized diet than modern omni-herbivorous black bears, with none of the dental evidence of hard food consumption (such as carcasses or nuts) found in polar bears, black bears and hyenas.[21] Comparisons with the dental microwear of Ursus speleaus suggest dietary differences between the species, with cave bears consuming tougher vegetation than A. simus.[92] Although some researchers argue that herbivory should be more obvious from the isotope data gathered from northern Arctodus,[57] several Arctoduscoprolites from The Mammoth Site in South Dakota and Meander Cave at Ni'iinlii'njik Territorial Park, Yukon contain Juniperus seeds (toxic to black & brown bears).[63][96]
Opportunistic carnivory
Arctodus' closest living relative is the spectacled bear. Although mostly herbivorous, Tremarctos ornatus is on occasion an active predator.
Evidence suggests that Arctodus also consumed meat, as evidenced by elevated nitrogen-15 isotope levels (corresponding to protein consumption) and bone damage on contemporary fauna. Additionally, elevated carbon-13 levels (corresponding to C3 resources) from many localities (Alaska,[98] California,[28][91]San Luis Potosí,[29] Texas,[99] Vancouver Island,[89][100] and the Yukon)[101] largely suggest browsers (and browsed vegetation) were the core of A. simus' diet.[44]
Arctodus simus' status as a predator is questioned by its gracility and lack of agility, which could have complicated predation upon adult mega-herbivores, and hindered the chasing down of nimbler prey.[72][74] Nevertheless, larger (male) Arctodus simus are suggested to have been more carnivorous than females, as very large brown bears may not be able to sustain themselves on a vegetarian diet.[100] Furthermore, the much larger frame of A. simus would have provided an advantage in disputes over carcasses.[19]
Studies establish that Arctodus simus would have had a varied diet across its range,[21] and was outcompeted and/or more herbivorous with increased competition from other predators.[85][100][98][91] The extinction of cursorial, hyper-carnivorous Borophagus and Huracan in the more open western North America left a vacant niche, possibly contributing to the evolution of A. simus (along with changes to the herbivore guild).[75][71]
Bone damage
Arctodus may have found young proboscideans to be suitable prey.
The bite marks found on many bones of ground sloths (Northrotheriops texanus) and young proboscideans at Leisey Shell Pit in Florida matched the size of the canine teeth of Arctodus pristinus. It is not known if these bite marks are the result of active predation or scavenging.[1] Additionally, Arctodus pristinus was the most common large predator from Port Kennedy Cave, Pennsylvania, where the majority of mastodon remains were juveniles and likely represent accumulated prey.[102]
Arctodus simus has been found in association with proboscidean remains near Frankstown, Pennsylvania (juvenile mastodon), and at The Mammoth Site, South Dakota (Columbian mammoths). However, questions remain as to whether these finds determine a predatory or scavenging relationship, or whether they were simply preserved at the same deposit (possibly at different times).[63][103] On the other hand, a woolly mammoth specimen from Saltville, Virginia was likely scavenged on by Arctodus simus, as evidenced by a canine gouge through the calcaneus.[57] Several Columbian mammoth bones from a cave near Huntington Reservoir, Utah also record ursid gnaw marks attributed to Arctodus, with an Arctodus specimen preserved in association with the remains.[61]
Importantly, the canines of Panthera atrox overlap in size with Arctodus simus, complicating the identification of tooth marks.[57] However, this is not to discredit all tooth marks attributed to Arctodus, as damaged bones from near Tanana River, Alaska suggest that Arctodus transported megafaunal longbones back to a cave-like den and chewed on them,[104][66][105] at a time when lions had a limited overlap with Arctodus in Beringia.[106][85] Furthermore, a perforated peccary ilium from Sheriden Cave has also been hypothesized as being scavenged by Arctodus simus.[107] Bone damage on a cranial fragment (and possibly the humerus) of an Arctodus individual on Vancouver Island may have been due to cannibalism.[17][100]
Beringia
Analysis of bones from Alaska showed high concentrations of nitrogen-15, a nitrogen isotope accumulated most strongly in carnivores. Although few specimens exist, there is currently no evidence of the same carbohydrate-related dental pathologies evident in southern populations of Arctodus simus.[85] Based on this evidence, A.simus was suggested to have been more carnivorous in Beringia than the rest of North America (with a preference for herbivores which consumed C3 vegetation, particularly caribou).[98][101][108] Increased carnivory may be due to a lower proportion of competitors and probably a lower availability of carbohydrate-rich food supplies across the year in the far northern latitudes.[85] Survival during the cold season for some northern populations of A. simus could have depended on the regular scavenging of ungulate carcasses, as is the case with Alaskan brown bears.[44][91] Ultimately, an opportunistic foraging strategy including up to 50% vegetation, and the meat of reindeer, muskox, carrion, and possibly some predators, is consistent with the isotopic data and the conclusions of the ecomorphological studies.[98]
Carbon isotope studies
Although elevated nitrogen-15 levels have been argued to indicate carnivory, even the isotope data of the most carnivorous Beringian Arctodus overlapped with modern, typically omni-herbivorous brown bears from Europe, eastern Wyoming, and central Montana, demonstrating that isotope data cannot distinguish between hypercarnivores and omnivores which eat a significant amount of animal matter.[46][98] Studies are also complicated by a lack of compound-specific data,[91] and isotope data being variable in carbon-13,[109][110] and nitrogen-15 (due to individual/evolving prey and plant choices, the isotopic composition of the local environment, and nutritional stress).[101][111]Carbon-13 levels in Arctodus simus (enriched by both plants and prey matter) consistently reflect a diet based on C3 resources, typically found in closed to mixed habitats with at least some tree cover (such as open woodlands).[28][29][89][98][91][99] This includes C3 vegetation (leaves, stems, fruits, bark, and flowers from trees, shrubs, and cool season grasses)[26] and the browsers that fed on them, such as deer, camelids, tapir, bison and ground sloths.[29][44]
Arctodus simus carbon isotope values from Irvington (along with Fairmead Landfill and McKittrick Tar Pits) are consistent with diet based on C3 resources.[28]
This Arctodus individual had the strongest δ13C value of its local fauna. Arctodus' carbon isotope value was closest to values from the tapir and Hemiauchenia.[29]
The Natural Trap specimens have the lowest δ13C of the Pleistocene fauna. Comparisons with contemporary Ursus suggest that the seasonality & individual choices within omnivorous diets could result in extreme isotope data, with each tooth not fully reflecting the total isotopic range consumed.[110]
Nitrogen isotope signatures suggested a ~19% consumption rate of seals (along with bison and camels). Fossil was likely transported post-mortem from the mainland; a partial reliance on marine resources has been suggested to be as a result of a competitive carnivore guild on mainland California. The marine signal was in between island foxes and bald eagles, most closely resembling Late Pleistocene California condors.[91]
Nitrogen & carbon isotope data from several specimens suggests that Arctodus specialized on reindeer in central Alaska, both before and during the Last Glacial Maximum.[101]
Arctodus'nitrogen-15 levels are higher in the Yukon, suggesting a possibly even higher trophic level than other Arctodus in eastern Beringia. However, this contrast likely reflects subtle differences in the isotopic composition of local plants,[59][113] & muskox in the region, and possibly fellow predators and their kills, complimenting the consumption of reindeer.[98]
Arctodus pristinus specimens have been found in caves such as Port Kennedy, Pennsylvania (where fossils from as many as 36 individuals have been found), and Cumberland Cave, Maryland, often in association with the black bear. This suggests a close association with the biome.[2][114]
According to a 2003 study, in karst regions, fossils of Arctodus simus have been recovered almost exclusively from cave sites.[54] In the contiguous United States, that ~38% of all sites are from caves (possibly ~50% in western USA)[51] suggests a close association between this species and cave environments. Cave passages produce numerous examples of small, yet relatively complete individuals. Taking into account that female bears are smaller and more prone to den in caves, it is likely that the majority of Arctodus simus from such deposits were females and may have been denning when they died.[54] Although female specimens of Arctodus simus have been inferred to have been exhibiting maternal denning, the expression of metabolic denning (hibernation/torpor) is unclear in Arctodus.[55] Moreover, to date, there are no records of adults with associated offspring from caves.[56] However, Arctotherium angustidens, a fellow giant short-faced bear, has been recovered from a cave in Argentina with offspring.[115]
At Riverbluff Cave, the most abundant claw marks are from Arctodus simus. They are most abundant at the bear beds and their associated passageways, indicating a close relationship with denning.[82] Numerous "bear" beds often preserve Arctodus simus and both Pleistocene and modern American black bears in association (U.a. amplidens and U. a. americanus)- such deposits have been found in Missouri, Oklahoma and Potter Creek Cave, California. These mixed deposits are assumed to have accumulated over time as individual bears (including Arctodus) died during winter sleep.[5][116][117] Furthermore, environmental DNA suggests that Arctodus and black bears shared a cave in Chiquihuite cave, Zacatecas.[16] At Labor-of-Love cave, Nevada, both American black bears and brown bears have been found in association with Arctodus simus. A study in 1985 noted that sympatry between Arctodus and brown bears preserved in caves is rare, with only Little Box Elder Cave, Wyoming and Fairbanks II, Alaska hosting similar remains.[26][48]
Although smaller than its descendant, Arctodus pristinus was still a relatively large tremarctine bear.[1] Sometimes referred to as the eastern short-faced bear,[118]A. pristinus has been found in Florida,[2] Kansas,[7] Maryland,[7] New Mexico,[119] Pennsylvania,[51][114] South Carolina,[120] and West Virginia in the US,[7] and Aguascalientes in Mexico.[121] Possible remains have also been recovered from Arizona.[7][122]A. pristinus is particularly well known from Florida, especially from the Leisey Shell Pit.[123] Like A. simus and other tremarctine bears, A. pristinus had adaptations for herbivory, and was likely largely herbivorous itself,[2] although Arctodus has been suggested to be generally more carnivorous than contemporary bears.[1][36]
Evolving from the smaller A. pristinus around 1.36 million years ago, scholars today mostly conclude that Arctodus simus was a colossal, opportunistic omnivore, with a flexible, locally adapted diet akin to the brown bear.[44][60][17][91][37] If Arctodus simus wasn't largely herbivorous,[2][26] the scavenging of megaherbivore carcasses, and the occasional predatory kill would have complimented the large amounts of vegetation consumed when available.[44][46][17][126]
Sometimes referred to as the bulldog bear,[130][131] or great short-faced bear,[15][132]Arctodus simus has been recovered from a comparatively small number of finds in relation to other large carnivorans, with the species suggested to have lived in low population densities.[16] However, A. simus was relatively plentiful in western North America, with over 50% of specimens from the contiguous United States (<40,000 BP).[31][85]Arctodus simus was integral to what has been referred to as the Camelops fauna, or alternatively Camelops/"Navahoceros" fauna, a faunal province centered in western North America. The Camelops fauna was also characterized by shrub-ox, prairie dogs, dwarf pronghorns, Shasta ground sloths, and American lions. The diverse flora of the Camelops faunal province included montane conifers and oak parklands, shrub and grassland that stretched across the North American Cordillera south of Canada, to the Valley of Mexico. This faunal province supported a variety of large grazing and browsing mammals.[27][133][134] Matheus argues that unlike other Holoarctic carnivorans, A. simus did not appear to have an ecological equivalent ("super-huge bear") in the Palearctic realm.[52]
Despite the shift to aridified environments from the Early to Late Pleistocene of the Central Valley (1.1Ma to ~15,000 BP), Arctodus simus remained consistent with the consumption of C3 resources. This period saw the evolution from wetter mixed woodland-grassland and marsh/prairie C3 dominated environs at Irvington and Fairmead, to the more arid, mixed C3-C4 savannas of the McKittrick Tar Pits. Dire wolves and Arctodus simus were consistent members of the local predator guild throughout the Pleistocene, whereas jaguars, Homotherium, Miracinonyx and Smilodon transitioned to Panthera atrox and coyotes.[28] In addition to browsing, Arctodus could have competed with other closed habitat browser specialists such as Smilodon and Panthera atrox for deer (Cervus & Odocoileus), camelids (Hemiauchenia & Camelops), Paramylodon, and peccaries.[28] However, specimens collected from the La Brea Tar Pits show evidence that A. simus preferred a herbivorous diet. A. simus is particularly famous from fossils found in the La Brea Tar Pits, with 33 individuals recovered (the most of any locality).[138][21][139] As only one juvenile has been found from La Brea, A. simus is suggested to have been solitary.[126] Many more finds come from across California,[7][5] Vancouver Island,[17][89] and Washington,[89] where the semi-arid woodland/scrub transitioned to forest-steppe,[140] and open grasslands/heath.[89]
Comparatively, the Rocky Mountain System had the fewest number of specimens of Arctodus simus in western North America.[31] However, one of the youngest dated Arctodus simus is from a cave near Huntington Reservoir, Utah, which sits at an elevation of 2,740m (~9,000ft). The central and southern Rocky Mountains may have acted as refugia for boreal parkland megafauna from the plateau such as Arctodus simus,[61][27] with the Huntington specimen being the only confirmed extinct megafauna dated to the Younger Dryas of the Great Basin.[141] Other remains have been found from Wyoming (such as Natural Trap Cave),[142][143] and Montana.[144][145]
Intermontane Plateaus
A reconstruction of Rancholabrean New Mexico (White Sands).
The Intermontane Plateaus had the highest number of Arctodus simus specimens south of the ice sheets.[31][129] The region has yielded some of the largest specimens of A. simus, including, what was once the largest specimen on record, from Salt Lake Valley, Utah.[50] In contrast with other parts of North America, the plateaus received more rainfall during the Late Pleistocene, as glacially cooled air collided with hot desert air. As a result, this greatly expanded the range of subalpine parkland, piñon-juniper & ponderosa woodlands, sagebrush grasslands and pluvial lakes where desert exists today.[141][140][51] The mid-Wisconsian U-Bar Cave, New Mexico, was vegetated by sagebrush, grasses, and woodlands. Notable fauna which lived alongside Arctodus simus included Shasta ground sloth, shrub-ox, pronghorns (Stockoceros,Capromeryx), Camelops, Odocoileus, horses, Lynx, puma, black bear, mountain goats, prairie dogs, and Stock's vampire bat.[30][146] Dire wolves were also found in association with Arctodus simus, and both species are the most common large carnivorans of Rancholabrean New Mexico.[51] Beyond Utah and New Mexico,[51][147][148][149][150][151] other important US specimens have also been found in Arizona,[7] eastern California,[152] Idaho,[7] Nevada,[153] and eastern Oregon.[154][155][156]
The Interior Plains were composed of temperate steppe grassland,[140] and among the specimens yielded from this region is one of the largest Arctodus simus currently on record, from the banks of the Kansas river.[163] The late Irvingtonian Doeden gravel pits in Montana preserves an open grassland habitat, with riparian woodlands, and likely some shrublands.[164]Arctodus simus co-existed with ground sloths (Megalonyx, Paramylodon), Pacific mastodon, camels, and Bootherium.[165][166][35] As bison were yet to migrate into North America, Columbian mammoths and horses dominated these early Illinoian grasslands.[167]
In the Rancholabrean age, Arctodus simus, grey wolves and coyotes were part of a predator guild throughout the great plains, and were joined by Columbian mammoths, camels, Hemiauchenia, and American pronghorns. While the northern plains aridified into cold steppe (e.g. Mammoth site, South Dakota),[168] the southern plains were a parkland with riparian hackberry forests, and large expanses of mixed grass prairie grasslands grading into wet meadows, with limited seasonality. In the south (Lubbock Lake, Texas), this fauna was joined by Smilodon, dire wolves, grey fox and red fox, preying upon prairie dogs, horses (Equus & Haringtonhippus), peccaries, Odocoileus, Capromeryx, Bison antiquus and Holmesina.[168][169] Beyond Texas,[170]Arctodus has also been found in Iowa,[171] Kansas,[7][172] Nebraska,[7] and southern Canada (Alberta & Saskatchewan),[173][174][175] which when unglaciated, would have formed a tundra ecosystem with an ice-free corridor to Beringia.[176]
To the south, the Interior Highlands had a very high density of Arctodus simus specimens (second only to the black bear),[31][85] due to the high rate of preservation in the cave-rich region. Sympatry between the two species is most apparent in Missouri- Arctodus simus has been found in association with black bears at Riverbluff, Bat and Big Bear caves.[180] Big Bear Cave preserves fossilized hair associated with Arctodus.[54] During the Last Glacial Maximum, both bears were joined by dire wolves, coyotes, jaguars, snowshoe hare, groundhogs and beavers at Bat Cave, which also records thousands of Platygonus remains. These fauna inhabited well-watered forest-grassland ecotone with a strong taiga influence, although the region did occasionally cycle through drier, grassier periods. These open woodlands were dominated by pines and spruce, and to a lesser extent by oaks.[181][182][183][184][185]
Eastern USA
Lake Rousseau, Florida, is the south-easternmost locality which Arctodus simus is known to have inhabited.
Compared to other regions, Arctodus simus was relatively rare in eastern North America.[19][31][85] To the north, the Appalachian Highlands were dominated by taiga.[140] Post-LGM Saltville, Virginia, was a mosaic of grassy/herb laden open areas intermixed with open canopy boreal woodlands (oaks, pines, spruce, birch, firs) and marshes. Inhabiting in this C3 resource dominated environment were Arctodus simus, mastodon, (southernmost) woolly mammoths, Bootherium, horses, caribou, Megalonyx, dire wolves, beavers, Cervalces, and a variety of warm-adapted reptiles, suggesting that a more mesic and less seasonal climate. Heavy bone damage on a mammoth carcass by both dire wolves and Arctodus suggests a potentially competitive scavenging relationship [186][57] Additional remains have been found in Virginia,[56] and Pennsylvania.[7][103]
To the south, the Atlantic Plains covered a great expanse of lowland, from the open deciduous woodlands of the Atlantic coast, to the semi-arid woodland/scrub of Florida, to the spruce-fir conifer forests and open habitat of the Gulf Coastal Plain. Although scarce, this contrast of habitats highlights the adaptability of Arctodus simus. At the Rainbow River and Lake Rousseau localities in Rancholabrean Florida, three Arctodus simus specimens have been recovered, alongside Smilodon, dire wolves, jaguars, ground sloths (Megalonyx, Paramylodon), llamas (Hemiauchenia, Palaeolama), Vero's tapir, giant beaver, capybara, Holmesina, horses, Bison antiquus, mastodon, Columbian mammoths and Tremarctos floridanus, in a climate similar to today's. Furthermore, the abundance of black bears, and particularly Tremarctos floridanus in Florida, has led to a theorized niche partitioning of ursids in Florida, with Tremarctos floridanus being herbivorous, and black bears and Arctodus simus being omnivorous, with Arctodus being possibly more inclined towards carnivory.[19] Additional finds of south-eastern Arctodus simus are from Alabama,[187] Arkansas,[188] Mississippi,[189][190][191] South Carolina,[192] and Texas.[62][193]
Mostly isolated by the Cordilleran and Laurentide ice sheets, Beringia is considered ecologically separate to the rest of North America, being largely an extension of the Eurasian mammoth steppe.[194] However, the occasional opening of an ice-free corridor, and the migration barrier of the Beringian gap, meant that Eastern Beringia (Alaska and the Yukon) supported a unique assemblage of fauna, with many endemic North American fauna flourishing.[73] This mostly open and treeless steppe-tundra, dominated by grasses, sedges, Artemisia spp., and a range of other forbs had a cold, dry climate, which prevented glaciation. Currently, all specimens of A. simus in Beringia have been dated to a 27,000 year window (50,000 BP - 23,000 BP) from Eastern Beringia,[106][17][7][53] while additional undated remains may be of Sangamonian age.[195][52] Unlike contemporary Beringian carnivorans, A. simus apparently never inhabited western Beringia (and therefore Asia).[52] The largest known skull of A. simus was recovered from the Yukon, and may represent the largest specimen known.[46][196]
The North Slope of Alaska <40,000 BP (Ikpikpuk and Titaluk rivers) preserves an upland and floodplain environment, with horses, bison then caribou being the most populous herbivores, and woolly mammoths, muskox, elk and saiga antelope more scarce. Cave lions, bears (Ursus arctos and Arctodus simus), and Beringian wolves made up the megafaunal predator guild.[197][198] Isotope data implies that caribou and muskox were principal components of the carnivorous portion of Arctodus simus' Arctic diet, suggesting that the warmer, wetter vegetation on the margins of the dry mammoth steppe (similar to the moist acidic tundra vegetation which dominates today) was the preferred habitat of Arctodus in Beringia.[98][197]
Additionally, upon the flooding of the Bering Strait and expansion of moist tundra and peatlands in Eastern Beringia during MIS-3, lions, brown bears and Homotherium went regionally extinct ~35,000 BP, whereas wolves and Arctodus persisted. Simultaneously, most megafaunal herbivores in Beringia experienced population bottlenecks, whilst mammoth populations steadily declined. This restriction of prey and habitat could explain the extinctions. However, genetically distinct cave lions and brown bears appear in MIS-2 circa the extinction of Arctodus in a re-emerged Beringia ~23,000 BP, opening up the possibility that some level of competition was at play.[106][98][199][200][201] The idea that Arctodus had a kleptoparasitic relationship with wolves and Homotherium in Beringia has been explored,[98] with the additional possibility that Arctodus successfully competed against brown bears and Homotherium for access to caribou pre-LGM.[101]
The local extinction of Arctodus in Beringia ~23,000 BP (possibly due to sharp climatic cooling associated with Heinrich Event-2),[106][17] was much earlier than in other parts of its range. While recolonized by cave lions and brown bears from Eurasia, Arctodus did not repopulate Beringia once the ice-free corridor to the south re-opened later in the Pleistocene.[106][202]
Radiocarbon dated A.simus (14C Date (1σ), ≤50,000 BP)
Relationships with other bears
Arctodus pristinus
In the Early Pleistocene, Arctodus pristinus was much more populous the south-east of North America, whereas the black bear was more common in the north-east.[203] The black bear has inhabited North America since at least the Middle Pleistocene,[100] while Tremarctos floridanus, a tremarctine bear inhabiting western North America at the time, is very similar to A. pristinus in terms of size, skeletal anatomy, and dietary preferences.[2]
Despite this, generally speaking large tremarctine fossils from the Early and Middle Pleistocene of Florida are considered to be A. pristinus, whereas those from the Late Pleistocene of Florida are considered to be T. floridanus. Indeed, black bears and Tremarctos floridanus are believed to have only colonized Florida with the extinction of A. pristinus (both of which only appear in Florida in the Late Pleistocene), however, T. floridanus could yet still be found from older sites in Florida.[2]T. floridanus was possibly being an ecological replacement of A. pristinus, with T. floridanus finds being widespread in Rancholabrean Florida and the wider southeastern United States.[2][23][31]
The evolution of Arctodus simus, competition with Tremarctos floridanus and black bears, and possibly the transitioning of PleistoceneFlorida from a hot, wet, densely forested habitat to a still hot, but drier and much more open biome are thought to be factors behind the gradual disappearance of Arctodus pristinus in the Middle Pleistocene (300,000 BP).[19][31] There are dubious records of A. pristinus in South Carolina and California from the Late Pleistocene,[67][204] however these are heavily disputed,[120][91] with modern research establishing A. pristinus existing between the Pliocene-Pleistocene boundary and the Middle Pleistocene.[2][19][120]
The most commonly accepted ecological parallel of Arctodus simus in scientific literature is the brown bear.[44][60][17] Both brown bears and Arctodus simus exhibit a high degree of dietary variability, and while largely herbivorous, meat can be an important dietary element to certain populations of both species.[98] Additionally, the potential of habitual kleptoparasitism is often noted in Arctodus, with brown bears being opportunistic, curious, and regularly stealing kills from smaller predators.[70][98] One past theory behind the extinction of Arctodus simus is that A. simus may have been out-competed by brown bears as the latter expanded southwards from eastern Beringia ~13,000 BP, and gradually established itself in North America.[48]
However this has been refuted as new dates establish an extended coexistence, with some isolated A. simus remains being re-evaluated as brown bears.[106][17] Brown bears (along with lions, bison and red foxes) first emigrated to North America via Beringia during the Illinoian Glaciation, with brown bears first arriving between ~177,000 BP and ~111,000 BP in East Beringia.[106] Genetic divergences suggest brown bears first migrated south during MIS-5 (~92,000 - 83,000 BP) upon the opening of the ice-free corridor,[106][100] with the first fossils being near Edmonton (26,000 BP).[17] On a continent-wide scale, although the brown bear and Arctodus simus were sympatric at times as brown bears spread through North America, Arctodus simus may typically have dominated competitive interactions, and displaced brown bears from specific localities.[17] Additionally, Arctodus' prolonged co-existence with black bears may have put significant constraints on the black bear's evolution.[15]
At the end of the Pleistocene, one reason brown bears persisted where Arctodus simus went extinct was because Arctodus may have been less flexible in adapting to new and rapidly changing environments that impacted the availability or quality of food and habitat.[17] Brown bears and Arctodus have been discovered together in Alaska (then Beringia) between 50,000 BP and 34,000 BP,[106] and in later Pleistocene deposits in Vancouver Island, California, Wyoming and Nevada.[7][17]
Beringia
Isotope values (δ13C and δ15N) in numerous BeringianArctodus simus specimens suggests A. simus usually occupied a higher trophic level compared with invading brown bears. While some Beringian brown bears consumed salmon as well as vegetation, reduced signatures of terrestrial meat consumption were mostly noted. On the other hand, data from Beringian specimens of Arctodus clustered much more tightly, and suggested that only terrestrial sources of meat were important for Beringian Arctodus.[59] The forcing of a smaller bear into a more herbivorous diet has been compared to the modern relationship between brown bears and American black bears.[70][98] Where they overlap, black bears take the lower trophic niche, with lower population densities, much smaller territorial ranges, and seasonal migrations.[100] That Arctodus simus (along with local climate change) may have excluded brown bears from Eastern Beringia from ~34,000 to ~23,000 BP further suggests that Arctodus may typically have been dominant over brown bears.[199][200] When Arctodus went extinct in Beringia ~23,000 BP, brown bears recolonized Beringia, but had more carnivorous diets than their Beringian kin pre ~34,000 BP. This bolsters the idea that these bears competed for similar resources and niches.[106][17] Extinction and repopulation is further evidenced by the high genetic (mitochondrial) diversity of Beringian brown bears in contrast with Beringian Arctodus simus. This contrast in genetic diversity has also been hypothesized to suggest that while female brown bears have a permanent home range, female Arctodus simus may not have (at least not to the same extent).[90][106]
The brown bear was a direct competitor of Arctodus simus.
Vancouver Island
Brown bears, black bears and Arctodus simus all co-existed on Vancouver Island once the island de-glaciated ~14,500 BP.[100][17] According to an isotope analysis, all three bears relied on terrestrial resources, with black bears occupied a distinctly lower trophic position in relation to the brown bear, with Arctodus holding an intermediate position. This may be an underestimate, as the Arctodus specimens from Vancouver Island are believed to be female; as per brown & black bears, female A. simus may have had a significant decrease in protein consumption compared with male A. simus when co-existing with brown bears. Additionally, an analysis of Arctodus' data suggested that when consuming protein, meat was preferred.[100] It is possible that the each species was specialized to different environmental settings, which vary greatly across small geographical areas on the mountainous island, although both Arctodus simus and brown bears appeared to have preferred more open habitats.[100]
Convergent evolution
Both giant short-faced bearsArctodus simus and Arctotherium angustidens reached huge body sizes, in an example of convergent evolution.[75] However, beyond gigantism, there are notable differences between the species. Not only did Arctotherium angustidens reach a higher maximum weight (an exceptional specimen was calculated at ~1,670 kilograms (3,680lb)), A. angustidens was a much more robust animal, in contrast with the gracileArctodus simus.[38] Excluding the exceptional specimen, Arctotherium angustidens had been calculated to a weight range between 1,200 kilograms (2,600lb) and 412 kilograms (908lb),[205][44] with the largest specimens of either species being said to be comparable to one another.[205][60] The panda-relative Agriotherium africanum has also been suggested to share ecomorphological convergences with Arctodus simus.[46] Together with great size, the two species converged on several adaptations, including a skull with a short broad rostrums, premasseteric fossa on the mandible, possible carnassial shears (P4 and m1), and long limbs (relative to body length). These features were also shared by other extinct bears (Agriotherium, Huracan & Arctotherium bonariensis).[46] However, while Agriotherium & Huracan have definitive adaptions for meat-heavy diets stemming from a running, predatory lifestyle, Arctodus simus lacks similar adaptations beyond proportionally longer limbs.[71]
Interactions with humans
The Clovis people are the first known culture to have interacted with Arctodus.
One documented interaction with Clovis people is present at the Lubbock Lake Landmark, Texas. A likely already deceased Arctodus simus was processed for subsistence (butchery marks indicated skinning, de-fleshing and disarticulation) and tool production, much in the same way as a mammoth carcass (~13,000 BP / 11,100 14C BP ).[206][207] Additionally, other remains of the Arctodus simus have been found in association with Paleo-Indian artifacts in Sheriden Cave, Ohio,[177][107][208] and Huntington Dam, Utah,[61] with an A. simusfootbone fragment from Spalding, Idaho also being charred.[209][210] The direct relationship between humans and some associated Arctodus remains has been debated.[211][212][213] Human hunting and butchery of large megafauna, particularly mammoths and mastodon, would likely have put people in competition with Arctodus simus. Defense against these large bears and the abandonment of carcasses are plausible outcomes,[17] along with the possible caching and disposal of carcass remains underwater to mask its odor from Arctodus.[214]
Migration barrier hypothesis
In the late 1980s, Val Geist hypothesized that "specialist, aggressive, competitive Rancholabrean fauna" such as Arctodus were a barrier for humans (along with other Siberian megafauna such as moose, grey wolves and brown bears) when migrating into North America (both Beringia and below the ice sheets).[215] Male A. simus were the largest and most powerful carnivorous land mammals in North America, with the potential specialization in obtaining and dominating distant and scarce resources. Humans in this hypothesis, though familiar with brown bears, would not have been able to avoid predation or effectively compete with Arctodus simus and other large Pleistocene North American carnivores, making human expansion difficult in Beringia and impossible south of the ice sheets.[17][70][52] However, this theory has never been accepted by anthropologists.[52] Paul Matheus argues that there were negligible ecological differences across the mammoth steppe, and that humans successfully competed against and even hunted territorial cave bears, cave hyenas, cave lions, leopards, tigers and wolves in Eurasia before reaching eastern Beringia, making the solitary Arctodus an unlikely impediment to expansion.[52] Indeed, new dates establish an extended co-existence of humans and megafauna such as Arctodus across North America.[216][217][218][219]
Humans migrated to North America via the Siberian mammoth steppe, arriving at Eastern Beringia (Alaska and the Yukon). However, the migration was halted at the North American Ice Sheet, which separated Beringia and southern North America for most of the Late Pleistocene.[220] Both humans and Arctodus are first dated to ~50,000 BP in Beringia, both from sites in the Yukon, and co-existed until Arctodus went extinct in Beringia ~23,000 BP during the Last Glacial Maximum. This co-existence continued through the regional extinction of other Beringian predators such as cave lions, brown bears and saber-tooth cats.[106] Important sites of pre-LGM human occupation in Beringia include Old Crow Flats & the Klondike,[221][222]Kuparuk River Valley,[223] & the Bluefish Caves.[224][225]
Contiguous North America
The human colonization of North America south of the ice sheets further disproves the idea that Arctodus was a migration barrier. The earliest universally accepted pre-Clovis site south of Beringia are the White Sands footprints in New Mexico, dated to ~22,000 cal. BP.[216] Other pre-LGM sites across the Americas, such as Chiquihuite Cave,[226][217]Valsequillo,[227] El Cedral,[228] Santa Elina,[218]Gault,[229] and Hartley Mammoth Site,[230] affirm that humans proliferated alongside megafauna (such as Arctodus) in southern North America for more than ten thousand years.[217][219][229][230][231] Humans were definitively widespread across the Americas by at least 15,000 BP.[17][219]
Extinction
Skeletal reconstruction of Arctodus simus.
Arctodus pristinus went extinct in the late Irvingtonian (300,000 years ago),[19] being last recorded from the Coleman 2A site, Florida.[232] With the extinction of Arctodus pristinus, Arctodus simus became the final representative of the genus. Arctodus simus went extinct around 12,800 years ago, and is one of the most recently dated megafauna to go extinct in North America, being reliably dated to within the Pleistocene-Holocene boundary (13,800 BP - 11,400 BP).[233][129][234] Both local and regionalized dietary flexibility has been a factor suggested for the species' longevity.[85]
Various factors, including the depletion in number of large herbivores,[47][43] the diminishing nutritional quality of plants during climate change, and competition with fellow omnivores (humans and brown bears) for food resources, have been suggested as the cause of Arctodus simus' extinction.[206] However, multiple studies put doubt on brown bears being culpable in Arctodus simus' extinction, with the brown bear being more of an ecological replacement that was more adaptable to change.[31][17][43] Moreover, there is no systematic evidence that humans hunted large extinct Pleistocene carnivores in North America, and no clear indication of direct human involvement in the extinction of Arctodus simus.[17] Additionally, dental wear evidence from Rancho La Brea does not suggest that food shortages were to blame for the demise large bodied carnivorans such as Arctodus simus.[21]
Climate change
Of the factors discussed, vegetation shifts in the latest Pleistocene may have been particularly unfavorable for Arctodus simus, due to a reduction of quality foraging for subsistence. For example, on Vancouver Island (~13,500 BP), vegetation changed rapidly from open woodlands with abundant lodgepole pine to increasingly closed forests with shade-tolerant spruce, mountain hemlock, and red alder. These changes, effective by ~12,450 BP, point toward cool and moist conditions during the Younger Dryas stadial. Closed forests continued to expand in the early Holocene. Even though Arctodus simus was not restricted to open areas and could inhabit in different environments, the timing of the regional shift from an open pine woodland habitat to a densely forested vegetation implies that these vegetation changes contributed to the local extinction of Arctodus simus, along with many other megafauna.[17]
Low genetic diversity
Low genetic diversity could have hastened Arctodus' extinction.
Arctodus simus had a very low level of genetic diversity from most sampled specimens, albeit a sample with a Beringian and temporal bias (<44,000 BP). A loss and/or replacement of mitochondrial DNA lineages before the Last Glacial Maximum, and decrease in population size from a previously genetically diverse population, has been noted in a variety of Eurasian and American Late Pleistocene megafauna.[90][235] That the individual from Sheriden Cave, Ohio was very closely related to Beringian specimens may further support this idea, as these populations had possibly been isolated from before the Last Glacial Maximum (tens of thousands of years).[90]
A lack of genetic diversity has been attributed to a reduced ability to adapt to environmental conditions. Small population sizes may be characteristic of tremarctine bears- the spectacled bear, while having low levels of genetic diversity, has no signs of a recent genetic bottleneck. However, brown bears had diverse, sympatric source populations in Eurasia, allowing for repopulations/reinvasions into the Americas. If Arctodus simus experienced genetic bottlenecks or local extinctions prior to the Last Glacial Maximum, Arctodus would have been unable to supplement their reduced genetic diversity with new migrants like the brown bear could, making them vulnerable to extinction.[90]
Last dates
The youngest date for Arctodus simus is circa 12,700 BP from Friesenhahn Cave, Texas, calibrated from 10,814 ± 55 radiocarbon years (14C BP). However, this date should be viewed with caution, as analyses suggest the collagen protein was degraded. A vertebra from Bonner Springs, Kansas, was dated to ca. 12,800 BP (based on 10,921 ± 50 radiocarbon years) from well preserved collagen. However, another radiocarbon date from a different laboratory on the same vertebra widens the possible age of the vertebra to between 9,510 and 11,021 14C BP (at 2σ). Nevertheless, a specimen from Huntington Dam, Utah was also dated to ca. 12,800 BP from two radiocarbon dates (10,870 ± 75 & 10,976 ± 40 14C BP) and is therefore considered reliable.[56][233]
One past proposal envisaged A.simus as a brutish predator that overwhelmed very large but slow megafauna with its great physical strength.[72] However, despite being very large, its limbs were too gracile for such an attack strategy,[72][74][52] significantly more gracile so than Arctotherium angustidens at that.[38]
Due to their long legs, an alternative hypothesis suggested by Björn Kurtén is that it may have hunted by running down Pleistocene herbivores such as wild horses and saiga antelopes, an idea that at one time earned it the name "running bear".[43][44][131] However, during pursuit of speedy game animals, the bear's sheer physical mass, inflexible spine and plantigrade gait would be a handicap; modern brown bears can run at the same speed but quickly tire and cannot keep up a chase for long. Correspondingly, although a 700kg (1,500lb)Arctodus may have been able to reach a maximum speed of 51 kilometres per hour (32mph), all modern bears have maximum speeds significantly lower than mass-based calculations for speed. As a result, paleontologist Paul Matheus suggests that Arctodus' top speed was 40–45km/h (25–28mph). Arctodus skeletons do not articulate in a way that would have allowed for quick turns – an ability required of any predator that survives by chasing down agile prey.[39][46][72] Proportionally taller legs, a short trunk, proximally elongated limbs, a stride which had little to no unsupported intervals, small and laterally-orientated eyes, and proportionally short canines ill-suited for spinal and tracheal attacks further complicated ambush hunting as a lifestyle for Arctodus.[26][46][71]
Furthermore, the lack of definitive predatory adaptions (such as the absence of laterally compressed canines, and carnassials built for crushing and grinding rather than shearing meat) puts doubt to any species-wide hyper-carnivorous interpretations of Arctodus.[44][76][46][69] The anatomical requirements for a large, cursorial, hyper-carnivorous bear are present in Huracan and Agriotherium, but not Arctodus.[71][236] Adaptations for predatory behavior are highly divergent in ursids versus other carnivorans, with features such as a short rostrum and long carnassials not being indicative of a predatory lifestyle in Arctodus.[69] Although the only living hyper-carnivorousursid, the polar bear, also lacks carnassial shears, the species' specialization on small prey and reliance on blubber (rather than coarser flesh) invalidates a comparison with Arctodus.[26][46][69] Therefore, carnivory was limited to the regular scavenging of carcasses and opportunistic hunting, as is the case with the modern brown bear.[43][44][46]
The idea that Arctodus was an obligate kleptoparasite was most notably proposed by Paul Matheus.[39] Under this model, A.simus was ill-equipped to be an active predator, having evolved as a specialized scavenger adapted to cover an extremely large home range in order to seek out broadly and unevenly distributed mega-mammal carcasses.[46] There would have been additional selective pressure for increased body size, so that Arctodus could procure and defend carcasses from other large carnivores, some of which were gregarious, or chase them from their kills and steal their food.[72] Matheus calculated that with a hyper-carnivorous diet, a 700kg (1,500lb) Beringian Arctodus would need to consume ~5,853 kilograms (12,904lb) of meat per year- the equivalent of 12 bison, 44.6 horses, or 2 woolly mammoths (adjusted for the non-edible portions of the body). Therefore, Arctodus would have had to obtain 100kg (220lb) of flesh/edible carrion every 6.25 days (16kg (35.3lb) per day).[39][70][237]
Furthermore, the short rostrum, resulting in increased out-forces of the jaw-closing muscles (temporalis and masseter), may have been an adaptation for cracking bones with their broad carnassials. Such use of the P4 and m1 teeth is supported by the heavy wear on these teeth in old individuals of Arctodus simus and Agriotherium (another giant bear).[46] Additionally, strengthened tooth enamel in Arctodus may have evolved to crack bone.[68] Moreover, at least in Beringia, the conservative growth strategies, long lives and low natural mortality rates of horses and mammoths should have provided somewhat evenly distributed carcasses throughout the year (unlike ruminants such as bison, whose mortality peaks in late winter to early spring).[70] Finally, that Arctodus and the cave hyena did not spread into Siberia and North America respectively suggests some form of competitive exclusion was at play.[63]
Rebuttal
Clues from Arctodus' dentition, such as the absence of molar damage associated with processing bone, dental cavities, and the lack of specialisation in the canines, discourages a hyper-carnivorous interpretation of Arctodus.
The kleptoparasite hypothesis has been repeatedly challenged. The short, broad rostrum of Arctodus is a characteristic also shared with the sun bear and the spectacled bear, which are both omnivorous.[44] Specialized scavengers like hyenas show distinctive patterns of molar damage from cracking bones. Based on lack of "bone-cracking" wear in specimens from Rancho La Brea, researchers in 2013 concluded that Arctodus simus was not a specialized scavenger. Of living bears, this population of A. simus showed the most similar tooth wear patterns to its closest living relative, the spectacled bear, which can have a highly varied diet ranging from omnivory to almost pure herbivory.[26][21]
Additionally, severe tooth crown fractures and alveolar infections were found in the South American giant short-faced bear (Arctotheriumangustidens). These were interpreted as evidence of feeding on hard materials (e.g. bones), which could tentatively indicate for these bears the regular scavenging of ungulate carcasses obtained through kleptoparasitism. However, such dental pathologies were not observed in various specimens of A. simus, other than the strong wear facets of old individuals.[44] Instead, recovered dental damage (incisor wear, dental calculus & cavities) is herbivorous in origin.[54][85][17] Moreover, researchers in 2015 reviewing links between canine breakage, microwear texture patterns and carnivorans from La Brea found that A. simus consumed foods softer yet tougher than black bears and polar bears, avoided hard/brittle foods such as bone, and reaffirmed affinities between A. simus and modern, largely herbivorous spectacled bears.[93] In addition to hyenas, many other fauna did not cross the Rancholabrean Beringian gap, such as the American badger, Bootherium and the woolly rhino).[238][239][240]
Furthermore, the relative lack of Arctodus remains at predator traps such as the La Brea Tar Pits, suggests that Arctodus did not regularly compete for carcasses.[85] Although La Brea has produced more Arctodus simus specimens than any other site, Arctodus represents only 1% of all carnivorans in the pits.[93] While more abundant than brown bears and black bears, Arctodus was calculated to its baseline continental abundance, contrasting with the overabundance of other large carnivorans.[241] A similar rate (~0.9%) of relative abundance was calculated for Arctodus compared to other megafauna at the Natural Trap Cave in Wyoming by 1993.[242] Additionally, isotope analyses of Beringian Arctodus specimens suggest that Arctodus had a low consumption rate of horses and mammoths in Beringia, despite those species making up ~50% of the available biomass in Beringia.[98] Further evidence comes from the evolution of brain size relative to body size- bears with high caloric diets and which do not exhibit dormancy showed a weak but significant correlation with bigger relative brain size. Arctodus simus plotted in between the likely hypercarnivorous Cephalogale, and the almost exclusively herbivorous Eurasian cave bear and Indarctos, suggesting omnivory.[243]
A mammoth is any species of the extinct elephantid genus Mammuthus. They lived from the late Miocene epoch into the Holocene about 4,000 years ago, and various species existed in Africa, Europe, Asia, and North America. Mammoths are distinguished from living elephants by their spirally twisted tusks and in at least some later species, the development of numerous adaptions to living in cold environments, including a thick layer of fur.
The cave bear is a prehistoric species of bear that lived in Europe and Asia during the Pleistocene and became extinct about 24,000 years ago during the Last Glacial Maximum.
Beringia is defined today as the land and maritime area bounded on the west by the Lena River in Russia; on the east by the Mackenzie River in Canada; on the north by 72 degrees north latitude in the Chukchi Sea; and on the south by the tip of the Kamchatka Peninsula. It includes the Chukchi Sea, the Bering Sea, the Bering Strait, the Chukchi and Kamchatka Peninsulas in Russia as well as Alaska in the United States and the Yukon in Canada.
The dire wolf is an extinct canine. The dire wolf lived in the Americas during the Late Pleistocene and Early Holocene epochs. The species was named in 1858, four years after the first specimen had been found. Two subspecies are recognized: Aenocyon dirus guildayi and Aenocyon dirus dirus. The largest collection of its fossils has been obtained from the Rancho La Brea Tar Pits in Los Angeles.
The steppe bison or steppe wisent is an extinct species of bison. It was widely distributed across the mammoth steppe, ranging from Western Europe to eastern Beringia in North America during the Late Pleistocene. It is ancestral to all North American bison, including ultimately modern American bison. Three chronological subspecies, Bison priscus priscus, Bison priscus mediator, and Bison priscus gigas, have been suggested.
Panthera spelaea, also known as the cave lion or steppe lion, is an extinct Panthera species that most likely evolved in Europe after the third Cromerian interglacial stage, less than 600,000 years ago. Genetic analysis of ancient DNA has revealed that while closely related, it was a distinct species genetically isolated from the modern lion occurring in Africa and Asia, with the genetic divergence between the two species variously estimated between 1.9 million and 600,000 years ago. It is closely related and probably ancestral to the American lion. The species ranged from Western Europe to eastern Beringia in North America, and was a prominent member of the mammoth steppe fauna. It became extinct about 13,000 years ago.
Panthera atrox, better known as the American lion, also called the North American lion, or American cave lion, is an extinct pantherine cat. Panthera atrox lived in North America during the Pleistocene epoch, from around 340,000 to 12,800 years ago. The species was initially described by American paleontologist Joseph Leidy in 1853 based on a fragmentary mandible (jawbone) from Mississippi; the species name ('atrox') means "savage" or "cruel". The status of the species is debated, with some mammalogists and paleontologists considering it a distinct species or a subspecies of Panthera leo, which contains living lions. However, novel genetic evidence has shown that it is instead a distinct species derived from the Eurasian cave or steppe lion, evolving after its geographic isolation in North America. Its fossils have been excavated from Alaska to Mexico. It was about 25% larger than the modern lion, making it one of the largest known felids.
During the Last Glacial Maximum, the mammoth steppe, also known as steppe-tundra, was once the Earth's most extensive biome. It stretched east-to-west, from the Iberian Peninsula in the west of Europe, across Eurasia to North America, through Beringia and Canada; from north-to-south, the steppe reached from the arctic islands southward to China. The mammoth steppe was cold and dry, and relatively featureless, though topography and geography varied considerably throughout. Some areas featured rivers which, through erosion, naturally created gorges, gulleys, or small glens. The continual glacial recession and advancement over millennia contributed more to the formation of larger valleys and different geographical features. Overall, however, the steppe is known to be flat and expansive grassland. The vegetation was dominated by palatable, high-productivity grasses, herbs and willow shrubs.
Hippidion is an extinct genus of equine that lived in South America from the Late Pliocene to the end of the Late Pleistocene (Lujanian), between 2.5 million and 11,000 years ago. They were one of two lineages of equines native to South America during the Pleistocene epoch, alongside Equus (Amerhippus) neogeus.
The Tremarctinae or short-faced bears is a subfamily of Ursidae that contains one living representative, the spectacled bear of South America, and several extinct species from four genera: the Florida spectacled bear, the North American giant short-faced bears Arctodus, the South American giant short-faced bear Arctotherium as well as Plionarctos(P. edensis and P. harroldorum), which is thought to be ancestral to the other three genera. Of these, the giant short-faced bears may have been the largest ever carnivorans in the Americas. The group is thought to have originated in eastern North America, and then invaded South America as part of the Great American Interchange. Most short-faced bears became extinct at the end of the Pleistocene.
Tremarctos floridanus, occasionally called the Florida spectacled bear, Florida cave bear, or rarely Florida short-faced bear, is an extinct species of bear in the family Ursidae, subfamily Tremarctinae. T. floridanus was widespread in the Southeastern United States during the Rancholabrean epoch, with scattered reports of fossils from other parts of North America and from earlier epochs.
The Late Pleistocene to the beginning of the Holocene saw numerous extinctions of predominantly megafaunal animal species, which resulted in a collapse in faunal density and diversity across the globe. The extinctions during the Late Pleistocene are differentiated from previous extinctions by the widespread absence of ecological succession to replace these extinct megafaunal species, and the regime shift of previously established faunal relationships and habitats as a consequence. The timing and severity of the extinctions varied by region and are thought to have been driven by varying combinations of human and climatic factors. Human impact on megafauna populations is thought to have been driven by hunting ("overkill"), as well as possibly environmental alteration. The relative importance of human vs climatic factors in the extinctions has been the subject of long-running controversy.
Arctotherium is an extinct genus of the Pleistocene short-faced bears endemic to Central and South America. Arctotherium migrated from North America to South America during the Great American Interchange, following the formation of the Isthmus of Panama during the late Pliocene. The genus consists of one early giant form, A. angustidens, and several succeeding smaller species, which were within the size range of modern bears. Arctotherium was adapted to open and mixed habitat. They are genetically closer to the spectacled bear, than to Arctodus of North America, implying the two extinct forms evolved large size in a convergent manner.
Saltville Archaeological Site SV-2 an apparent Pre-Clovis archaeological site located in the Saltville Valley near Saltville, Virginia. The site was excavated from 1992 to 1997 by paleogeographer Jerry N. McDonald of the Virginia Museum of Natural History.
Valerius Geist was a Canadian biologist and a professor emeritus in the Faculty of Environmental Design at the University of Calgary. He was a specialist on the biology, behavior, and social dynamics of North American large mammals, and well respected on his views of Neanderthal people and behavior.
The Beringian wolf is an extinct population of wolf that lived during the Ice Age. It inhabited what is now modern-day Alaska, Yukon, and northern British Columbia. Some of these wolves survived well into the Holocene. The Beringian wolf is an ecomorph of the gray wolf and has been comprehensively studied using a range of scientific techniques, yielding new information on their prey species and feeding behaviors. It has been determined that these wolves are morphologically distinct from modern North American wolves and genetically basal to most modern and extinct wolves. The Beringian wolf has not been assigned a subspecies classification and its relationship with the extinct European cave wolf is not clear.
The Pleistocene wolf, also referred to as the Late Pleistocene wolf, is an extinct lineage or ecomorph of the grey wolf. It was a Late Pleistocene 129 Ka – early Holocene 11 Ka hypercarnivore. While comparable in size to a large modern grey wolf, it possessed a shorter, broader palate with large carnassial teeth relative to its overall skull size, allowing it to prey and scavenge on Pleistocene megafauna. Such an adaptation is an example of phenotypic plasticity. It was once distributed across the northern Holarctic. Phylogenetic evidence indicates that despite being much smaller than this prehistoric wolf, the Japanese wolf, which went extinct in the early 20th century, was of a Pleistocene wolf lineage, thus extending its survival to several millennia after its previous estimated extinction around 7,500 years ago.
Ursus ingressus is an extinct species of the family Ursidae that lived in Central Europe during the Late Pleistocene. It is named after the Gamssulzen Cave in Austria, where the holotype of this species was found.
The coastal migration hypothesis is one of two leading hypotheses about the settlement of the Americas at the time of the Last Glacial Maximum. It proposes one or more migration routes involving watercraft, via the Kurile island chain, along the coast of Beringia and the archipelagos off the Alaskan-British Columbian coast, continuing down the coast to Central and South America. The alternative is the hypothesis solely by interior routes, which assumes migration along an ice-free corridor between the Laurentide and Cordilleran ice sheets during the Last Glacial Maximum.
Panthera balamoides is a species described as an extinct species of the big cat genus Panthera that is known from a single fossil found in a Late Pleistocene age cenote in the Yucatan Peninsula, Mexico. P. balamoides has only a single reported specimen, the distal end of a right humerus, that is notably of exceptional size for a felid. It was unearthed in 2012 from an underwater cave and described in 2019 by an international group of paleontologists from Mexico and Germany led by Sarah R. Stinnesbeck. However, several authors have since proposed that the fossil comes from an ursid, possibly the extinct Arctotherium, and not of felid affinities.
1 2 3 Russell, Dale A.; Rich, Fredrick J.; Schneider, Vincent; Lynch-Stieglitz, Jean (May 2009). "A warm thermal enclave in the Late Pleistocene of the South-eastern United States". Biological Reviews. 84 (2): 173–202. doi:10.1111/j.1469-185X.2008.00069.x. PMID19391200. S2CID9609391.
1 2 Bell, Christopher; Lundelius, Ernest L.; Barnosky, Anthony D.; Zarzewski, Richard J.; Graham, Russell; Lindsay, Everett H.; Ruez, Dennis R.; Semken, Holmes A.; Webb, S. David (2004-04-21). "Chapter 7: The Blancan, Irvingtonian, and Rancholabrean Mammal Ages". In Woodburne, Michael O. (ed.). Late Cretaceous and Cenozoic Mammals of North America. Columbia University Press. doi:10.7312/wood13040. ISBN978-0-231-50378-5.
1 2 3 4 Soibelzon, Leopoldo H.; Schubert, Blaine W. (2011). "The Largest Known Bear, Arctotherium Angustidens, from the Early Pleistocene Pampean Region of Argentina: With a Discussion of Size and Diet Trends in Bears". Journal of Paleontology. 85 (1): 69–75. doi:10.1666/10-037.1. hdl:11336/104215. JSTOR23019499. S2CID129585554.
1 2 Nelson, Michael E.; Madsen, James H. (1983). "A Giant Short-Faced Bear (Arctodus simus) from the Pleistocene of Northern Utah". Transactions of the Kansas Academy of Science. 86 (1): 1–9. doi:10.2307/3628418. JSTOR3628418.
1 2 Fowler, Nicholas L.; Spady, Thomas J.; Wang, Guiming; Leopold, Bruce D.; Belant, Jerrold L. (October 2021). "Denning, metabolic suppression, and the realisation of ecological opportunities in Ursidae". Mammal Review. 51 (4): 465–481. doi:10.1111/mam.12246. S2CID233847639.
1 2 3 4 Schubert, Blaine W. (2010-04-15). "Late Quaternary chronology and extinction of North American giant short-faced bears (Arctodus simus)". Quaternary International. Faunal Dynamics and Extinction in the Quaternary: Studies in Honor of Ernest L. Lundelius, Jr. 217 (1): 188–194. Bibcode:2010QuInt.217..188S. doi:10.1016/j.quaint.2009.11.010.
1 2 3 4 Gillette, David D.; Madsen, David B. (1992-03-06). "The short-faced bear Arctodus simus from the late Quaternary in the Wasatch Mountains of central Utah". Journal of Vertebrate Paleontology. 12 (1): 107–112. Bibcode:1992JVPal..12..107G. doi:10.1080/02724634.1992.10011436.
1 2 3 Slaughter, B. H.; Crook, W. W. Jr.; Harris, R. K.; Allen, D. C.; Seifert, M. (1962-01-01). The Hill-Shuler Local Faunas of the Upper Trinity River, Dallas and Denton Counties, Texas. Report Investigation. The University of Texas at Austin, Bureau of Economic Geology. doi:10.23867/ri0048d.
1 2 Voorhies, M. R.; Corner, R. G. (1986). "Giant Bear Arctodus as a Potential Breaker and Flaker of Late Pleistocene Megafaunal Remains". Current Research in the Pleistocene. 3: 49–51.
1 2 3 4 5 6 7 8 Matheus, Paul E. (2003). Locomotor adaptations and ecomorphology of short-faced bears (Arctodus simus) in eastern Beringia. Yukon Palaeontologist, Gov't. of Yukon. OCLC243520303.
1 2 3 Lynch, Eric Randally (2012-08-06). Cursorial Adaptations in the Forelimb of the Giant Short-Faced Bear, Arctodus simus, Revealed by Traditional and 3D Landmark Morphometrics. East Tennessee State University. OCLC818344518.
↑ Stucchi, Marcelo; Salas-Gismondi, Rodolfo; Baby, Patrice; Guyot, Jean-Loup; Shockey, Bruce J. (2009). "A 6,000+ year-old specimen of a spectacled bear from an Andean cave in Peru". Ursus. 20: 63–68. doi:10.2192/08GR017R1.1. S2CID86731722.
↑ Nadeau, Andy J.; Allen, Kathy; Benck, Kevin; Davis, Anna M.; Hutchins, Hannah; Gardner, Sarah; Amberg, Shannon; Robertson, Andrew (September 2017). "White Sands National Monument Natural Resource Condition Assessment"(PDF). Natural Resource Report NPS/WHSA/NRR—2017/1508: 276–277 – via National Park Service, Fort Collins, Colorado.
1 2 Lucas, Spencer G.; Spielmann, Justin A.; Lockley, Martin G. (2007). "Cenozoic Vertebrate Tracks and Traces". New Mexico Museum of Natural History and Science Bulletin. 42.
↑ Rovey II, Charles W.; Forir, Matt; Balco, Greg; Gaunt, David (2010-01-01). "Geomorphology and Paleontology of Riverbluff Cave, Springfield, Missouri". In Evans, Kevin R.; Aber, James S. (eds.). From Precambrian Rift Volcanoes to the Mississippian Shelf Margin: Geological Field Excursions in the Ozark Mountains (Field Guide 17ed.). Geological Society of America. pp.31–32. ISBN978-0-8137-0017-5.
1 2 3 4 5 6 7 8 9 10 11 12 13 Salis, Alexander T; Bray, Sarah C E; Lee, Michael S Y; Heiniger, Holly; Barnett, Ross; Burns, James A; Doronichev, Vladimir; Fedje, Daryl; Golovanova, Liubov; Harington, C Richard; Hockett, Bryan; Kosintsev, Pavel; Lai, Xulong; Mackie, Quentin; Vasiliev, Sergei; Weinstock, Jacobo; Yamaguchi, Nobuyuki; Meachen, Julie; Cooper, Alan; Mitchell, Kieren J (3 September 2020). "Lions and brown bears colonized North America in multiple synchronous waves of dispersal across the Bering Land Bridge". bioRxiv10.1101/2020.09.03.279117.
↑ Puckette, William L. (1976). "Notes on the occurrence of the short-faced bear (Arctodus) in Oklahoma". Proceedings of the Oklahoma Academy of Science. 56: 67–68. CiteSeerX10.1.1.605.3584.
↑ Grumet, Robert S. (September 2000). Bay, Plain, and Piedmont- A Landscape History of the Chesapeake Heartland from 1.3 Billion Years Ago to 2000. The Chesapeake Bay Heritage Context Project. pp.16, 21, 167.
↑ Dalquest, W. W.; Mooser, O. (1980-12-19). "Arctodus pristinus Leidy in the Pleistocene of Aguascalientes, Mexico". Journal of Mammalogy. 61 (4): 724–725. doi:10.2307/1380320. JSTOR1380320.
↑ Dundas, Robert G.; Chatters, James C. (2013-01-01). "The mid-Irvingtonian Fairmead Landfill fossil site, Madera County Paleontology Collection, and Fossil Discovery Center of Madera County, California". In Keith Putirka (ed.). Geologic Excursions from Fresno, California, and the Central Valley: A Tour of California's Iconic Geology. Geological Society of America. pp.63–78. doi:10.1130/2013.0032(04). ISBN978-0-8137-0032-8.
↑ Cassiliano, Michael L. (1999). "Biostratigraphy of Blancan and Irvingtonian Mammals in the Fish Creek-Vallecito Creek Section, Southern California, and a Review of the Blancan-Irvingtonian Boundary". Journal of Vertebrate Paleontology. 19 (1): 169–186. doi:10.1080/02724634.1999.10011131. ISSN0272-4634. JSTOR4523978.
↑ Long, C. A. (1971). "Significance of the Late Pleistocene fauna from the Little Box Elder Cave, Wyoming, to studies of zoogeography of recent mammals". Great Basin Naturalist. 31 (2) 11. S2CID55933331.
↑ Schultz, C. Bertrand; Howard, Edgar B.; Schultz, C. Bernard (1935). "The Fauna of Burnet Cave, Guadalupe Mountains, New Mexico". Proceedings of the Academy of Natural Sciences of Philadelphia. 87: 273–298. ISSN0097-3157. JSTOR4064215.
↑ Harris, Arthur H. (1993). "Quaternary Vertebrates of New Mexico"(PDF). Vertebrate Paleontology in New Mexico, New Mexico Museum of Natural History and Science. Bulletin 2: 179–197.
↑ Morgan, Gary S.; Lucas, Spencer G.; Love, David (2009). "Cenozoic vertebrates from Socorro County, central New Mexico"(PDF). In Virgil Lueth; Spencer G. Lucas; Richard M. Chamberlin (eds.). New Mexico Geological Society Fall Field Conference Guidebook: 60 Geology of the Chupadera Mesa. pp.321–336.
↑ Carranza-Castañeda, Oscar; Miller, Wade E. (16 September 1987). "Rediscovered type specimens and other important published Pleistocene mammalian fossils from Central Mexico". Journal of Vertebrate Paleontology. 7 (3): 335–341. Bibcode:1987JVPal...7..335C. doi:10.1080/02724634.1987.10011664.
↑ Young, Robert R.; Burns, James A.; Smith, Derald G.; Arnold, L. David; Rains, R. Bruce (1994-08-01). "A single, late Wisconsin, Laurentide glaciation, Edmonton area and southwestern Alberta 2.3.CO;2". Geology. 22 (8): 683–686. doi:10.1130/0091-7613(1994)022<0683:ASLWLG>2.3.CO;2. ISSN0091-7613.
↑ Wilson, Ronald C. (1985). "Vertebrate Remains in Kentucky Caves". In Percy H. Dougherty (ed.). Caves and Karst of Kentucky(PDF). Series XI. Vol.12. Kentucky Geological Survey, University of Kentucky Lexington. p.171. ISSN0075-5613.
↑ Slaughter, Bob H. (1966). "The Moore Pit Local Fauna; Pleistocene of Texas". Journal of Paleontology. 40 (1): 78–91. ISSN0022-3360. JSTOR1301775.
↑ David Webb, S.; Graham, Russell W.; Barnosky, Anthony D.; Bell, Christopher J.; Franz, Richard; Hadly, Elizabeth A.; Lundelius, Ernest L.; Gregory McDonald, H.; Martin, Robert A. (2003), Vertebrate paleontology, Developments in Quaternary Sciences, vol.1, Elsevier, pp.519–538, doi:10.1016/s1571-0866(03)01025-x, ISBN978-0-444-51470-7, retrieved 2022-06-28
↑ Holliday, Vance T.; Johnson, Eileen; Haas, Herbert; Stuckenrath, Robert (1983). "Radiocarbon Ages from the Lubbock Lake Site, 1950-1980: Framework for Cultural and Ecological Change on the Southern High Plains". Plains Anthropologist. 28 (101): 165–182. doi:10.1080/2052546.1983.11909122. ISSN0032-0447. JSTOR25668371.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.