
Chondrichthyes is a class of jawed fish that contains the cartilaginous fish or chondrichthyians, which all have skeletons primarily composed of cartilage. They can be contrasted with the Osteichthyes or bony fish, which have skeletons primarily composed of bone tissue. Chondrichthyes are aquatic vertebrates with paired fins, paired nares, placoid scales, conus arteriosus in the heart, and a lack of opecula and swim bladders. Within the infraphylum Gnathostomata, cartilaginous fishes are distinct from all other jawed vertebrates.

Chimaeras are cartilaginous fish in the order Chimaeriformes, known informally as ghost sharks, rat fish, spookfish, or rabbit fish; the last three names are not to be confused with rattails, Opisthoproctidae, or Siganidae, respectively.

Elasmobranchii is a subclass of Chondrichthyes or cartilaginous fish, including modern sharks, rays, skates, and sawfish. Members of this subclass are characterised by having five to seven pairs of gill clefts opening individually to the exterior, rigid dorsal fins and small placoid scales on the skin. The teeth are in several series; the upper jaw is not fused to the cranium, and the lower jaw is articulated with the upper. The details of this jaw anatomy vary between species, and help distinguish the different elasmobranch clades. The pelvic fins in males are modified to create claspers for the transfer of sperm. There is no swim bladder; instead, these fish maintain buoyancy with large livers rich in oil.

Acanthodii or acanthodians is an extinct class of gnathostomes. They are currently considered to represent a paraphyletic grade of various fish lineages basal to extant Chondrichthyes, which includes living sharks, rays, and chimaeras. Acanthodians possess a mosaic of features shared with both osteichthyans and chondrichthyans. In general body shape, they were similar to modern sharks, but their epidermis was covered with tiny rhomboid platelets like the scales of holosteians.

Holocephali, sometimes given the term Euchondrocephali, is a subclass of cartilaginous fish in the class Chondrichthyes. The earliest fossils are of teeth and come from the Devonian period. Little is known about these primitive forms, and the only surviving group in the subclass is the order Chimaeriformes.

Helicoprion is an extinct genus of shark-like eugeneodont fish. Almost all fossil specimens are of spirally arranged clusters of the individuals' teeth, called "tooth whorls", which in life were embedded in the lower jaw. As with most extinct cartilaginous fish, the skeleton is mostly unknown. Fossils of Helicoprion are known from a 20 million year timespan during the Permian period from the Artinskian stage of the Cisuralian to the Roadian stage of the Guadalupian. The closest living relatives of Helicoprion are the chimaeras, though their relationship is very distant. The unusual tooth arrangement is thought to have been an adaption for feeding on soft bodied prey, and may have functioned as a deshelling mechanism for hard bodied cephalopods such as nautiloids and ammonoids. In 2013, systematic revision of Helicoprion via morphometric analysis of the tooth whorls found only H. davisii, H. bessonowi and H. ergassaminon to be valid, with some of the larger tooth whorls being outliers.

Stethacanthus is an extinct genus of shark-like holocephalians which lived from the Late Devonian to Late Carboniferous epoch, dying out around 298.9 million years ago. Fossils have been found in Australia, Asia, Europe and North America.

Xenacanthida is an order or superorder of extinct shark-like chondrichthyans known from the Carboniferous to Triassic. They were native to freshwater, marginal marine and shallow marine habitats. Some xenacanths may have grown to lengths of 5 m (16 ft). Most xenacanths died out at the end of the Permian in the End-Permian Mass Extinction, with only a few forms surviving into the Triassic.

Hybodus is an extinct genus of hybodont that lived during the Early Jurassic. Species closely related to the type species Hybodus reticulatus lived during the Early Jurassic epoch. Numerous species have been assigned to Hybodus spanning a large period of time, and it is currently considered a wastebasket taxon that is 'broadly polyphyletic' and requires reexamination.

Stethacanthidae is an extinct family of prehistoric holocephalians. It is estimated to have existed approximately between 380 and 300 million years ago. Members of this family are noted for their peculiar dorsal fin.

Hybodontiformes, commonly called hybodonts, are an extinct group of shark-like cartilaginous fish (chondrichthyans) which existed from the late Devonian to the Late Cretaceous. Hybodonts share a close common ancestry with modern sharks and rays (Neoselachii) as part of the clade Euselachii. They are distinguished from other chondrichthyans by their distinctive fin spines and cephalic spines present on the heads of males. An ecologically diverse group, they were abundant in marine and freshwater environments during the late Paleozoic and early Mesozoic, but were rare in open marine environments by the end of the Jurassic, having been largely replaced by modern sharks, though they were still common in freshwater and marginal marine habitats. They survived until the end of the Cretaceous, before going extinct.
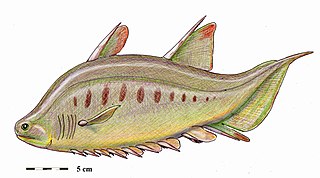
Brochoadmones is an extinct genus of acanthodian from the Devonian of what is now Canada. It is the only genus in the suborder Brochoadmonoidei, whose relationship to other acanthodian orders remains currently in flux.

Gyracanthides is an extinct genus of acanthodian gnathostome, known from Devonian to Early Carboniferous.
Dwykaselachus is an extinct genus of symmoriiform, a cartilaginous fish that lived in what is now South Africa during the Permian period around 280 million years ago. It was first discovered in the 1980s, in a nodule of sediments from the Karoo Supergroup. Dwykaselachus was named based on Dwyka Group, the group of sedimentary geological formation in the southeastern part of Africa. It represents the place where the type species Dwykaselachus oosthuizeni was found.

Plesioselachus is an extinct genus of Late Devonian (Famennian) cartilaginous fish with uncertain classification, which contains only one species, P. macracanthus from the Waterloo Farm lagerstätte in South Africa. Known from a single incomplete articulated skeleton and some isolated remains, it is characterized by having a long dorsal spine with length about one third of body length.

Synechodus is an extinct genus of shark belonging to the family Palaeospinacidae and order Synechodontiformes. It is known from 16 species primarily spanning from the Late Triassic to Paleocene. The dentition is multicusped and was used for grasping. Several species are known from skeletal remains, including the species Synechodus ungeri from the Late Jurassic of Germany, which shows that it was relatively short with large pectoral fins and a proportionally large head with a round snout. This species is suggested to have reached a body length of 30–40 centimetres (0.98–1.31 ft). Skeletal remains are also known of the species Synechodus dubrisiensis from the Cretaceous of Europe. A skeleton of an indeterminate species is also known from the Early Cretaceous (Albian) of France, with a body length of around 70 centimetres (2.3 ft). Synechodus is suggested to have had two dorsal fins that lacked fin spines, though the number of dorsal fins is unknown in Synechodus ungeri.

Ctenacanthiformes is an extinct order of cartilaginous fish. They possessed ornamented fin spines at the front of their dorsal fins and cladodont-type dentition, that is typically of a grasping morphology, though some taxa developed cutting and gouging tooth morphologies. Some ctenacanths are thought to have reached sizes comparable to the great white shark, with body lengths of up to 7 metres (23 ft) and weights of 1,500–2,500 kilograms (3,300–5,500 lb). The earliest ctenacanths appeared during the Frasnian stage of the Late Devonian, with the group reaching their greatest diversity during the Early Carboniferous (Mississippian), and continued to exist into at least the Middle Permian (Guadalupian). Some authors have suggested members of the family Ctenacanthidae may have survived into the Cretaceous based on teeth found in deep water deposits of Valanginian age in France and Austria, however, other authors contend that the similarity of these teeth to Paleozoic ctenacanths is only superficial, and they likely belong to neoselachians instead.

Squatinactis is a genus of extinct elasmobranch chondrichthyan known from the Carboniferous aged Bear Gulch Limestone in Montana. This fish was discovered in 1974 by Richard Lund. The type specimen, named CMNH 46133, consists of a brain case, poorly preserved jaws and gills, a pectoral fin, and a partial vertebral axis. This creatures most startling feature were its broad pectoral fins which resembled those of stingrays and angel sharks (Squatina). The holotype specimen has about 15 teeth in its jaw. This creature is named after the angel shark. Remains found in the South Urals of Russia and the Eyam Limestone of Derbyshire, England, have been tentatively identified as those belonging to S. caudispinatus.
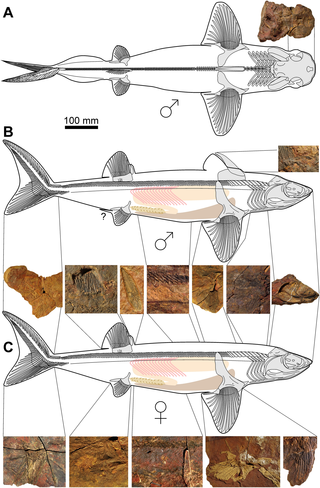
Maghriboselache is an extinct genus of cladoselachid symmoriiform fish that lived 369 million years ago during the Late Devonian of Morocco. The genus contains a single species, M. mohmezanei. Maghriboselache represents the first cladoselachid to be discovered with significant details of the jaws, braincase and even endocranium. Along with Cladoselache, it shares a unique and distinctive tooth and upper jaw morphology.
Cosmoselachus is an extinct genus of symmoriiform chondrichthyan from the Upper Carboniferous aged Fayetteville Shale of Arkansas, United States. The genus contains a single species, C. mehlingi, which is known from a partial specimen that includes the cranium, jaws, gill arches, pectoral fins, and teeth.

















