Related Research Articles

An oncogene is a gene that has the potential to cause cancer. In tumor cells, these genes are often mutated, or expressed at high levels.

In biochemistry, a kinase is an enzyme that catalyzes the transfer of phosphate groups from high-energy, phosphate-donating molecules to specific substrates. This process is known as phosphorylation, where the high-energy ATP molecule donates a phosphate group to the substrate molecule. This transesterification produces a phosphorylated substrate and ADP. Conversely, it is referred to as dephosphorylation when the phosphorylated substrate donates a phosphate group and ADP gains a phosphate group. These two processes, phosphorylation and dephosphorylation, occur four times during glycolysis.

Paracrine signaling is a form of cell signaling, a type of cellular communication in which a cell produces a signal to induce changes in nearby cells, altering the behaviour of those cells. Signaling molecules known as paracrine factors diffuse over a relatively short distance, as opposed to cell signaling by endocrine factors, hormones which travel considerably longer distances via the circulatory system; juxtacrine interactions; and autocrine signaling. Cells that produce paracrine factors secrete them into the immediate extracellular environment. Factors then travel to nearby cells in which the gradient of factor received determines the outcome. However, the exact distance that paracrine factors can travel is not certain.
A mitogen-activated protein kinase is a type of protein kinase that is specific to the amino acids serine and threonine. MAPKs are involved in directing cellular responses to a diverse array of stimuli, such as mitogens, osmotic stress, heat shock and proinflammatory cytokines. They regulate cell functions including proliferation, gene expression, differentiation, mitosis, cell survival, and apoptosis.

The restriction point (R), also known as the Start or G1/S checkpoint, is a cell cycle checkpoint in the G1 phase of the animal cell cycle at which the cell becomes "committed" to the cell cycle, and after which extracellular signals are no longer required to stimulate proliferation. The defining biochemical feature of the restriction point is the activation of G1/S- and S-phase cyclin-CDK complexes, which in turn phosphorylate proteins that initiate DNA replication, centrosome duplication, and other early cell cycle events. It is one of three main cell cycle checkpoints, the other two being the G2-M DNA damage checkpoint and the spindle checkpoint.

Nerve growth factor (NGF) is a neurotrophic factor and neuropeptide primarily involved in the regulation of growth, maintenance, proliferation, and survival of certain target neurons. It is perhaps the prototypical growth factor, in that it was one of the first to be described. Since it was first isolated by Nobel Laureates Rita Levi-Montalcini and Stanley Cohen in 1956, numerous biological processes involving NGF have been identified, two of them being the survival of pancreatic beta cells and the regulation of the immune system.
Biological crosstalk refers to instances in which one or more components of one signal transduction pathway affects another. This can be achieved through a number of ways with the most common form being crosstalk between proteins of signaling cascades. In these signal transduction pathways, there are often shared components that can interact with either pathway. A more complex instance of crosstalk can be observed with transmembrane crosstalk between the extracellular matrix (ECM) and the cytoskeleton.
Mitogen Activated Protein (MAP) kinase kinase kinase, MAPKKK is a serine/threonine-specific protein kinase which acts upon MAP kinase kinase. Subsequently, MAP kinase kinase activates MAP kinase. Several types of MAPKKK can exist but are mainly characterized by the MAP kinases they activate. MAPKKKs are stimulated by a large range of stimuli, primarily environmental and intracellular stressors. MAPKKK is responsible for various cell functions such as cell proliferation, cell differentiation, and apoptosis. The duration and intensity of signals determine which pathway ensues. Additionally, the use of protein scaffolds helps to place the MAPKKK in close proximity with its substrate to allow for a reaction. Lastly, because MAPKKK is involved in a series of several pathways, it has been used as a therapeutic target for cancer, amyloidosis, and neurodegenerative diseases. In humans, there are at least 19 genes which encode MAP kinase kinase kinases:
The MAPK/ERK pathway is a chain of proteins in the cell that communicates a signal from a receptor on the surface of the cell to the DNA in the nucleus of the cell.

Platelet-derived growth factor receptors (PDGF-R) are cell surface tyrosine kinase receptors for members of the platelet-derived growth factor (PDGF) family. PDGF subunits -A and -B are important factors regulating cell proliferation, cellular differentiation, cell growth, development and many diseases including cancer. There are two forms of the PDGF-R, alpha and beta each encoded by a different gene. Depending on which growth factor is bound, PDGF-R homo- or heterodimerizes.
In molecular biology, extracellular signal-regulated kinases (ERKs) or classical MAP kinases are widely expressed protein kinase intracellular signalling molecules that are involved in functions including the regulation of meiosis, mitosis, and postmitotic functions in differentiated cells. Many different stimuli, including growth factors, cytokines, virus infection, ligands for heterotrimeric G protein-coupled receptors, transforming agents, and carcinogens, activate the ERK pathway.

Cyclin D is a member of the cyclin protein family that is involved in regulating cell cycle progression. The synthesis of cyclin D is initiated during G1 and drives the G1/S phase transition. Cyclin D protein is anywhere from 155 to 477 amino acids in length.
The ErbB family of proteins contains four receptor tyrosine kinases, structurally related to the epidermal growth factor receptor (EGFR), its first discovered member. In humans, the family includes Her1, Her2, Her3 (ErbB3), and Her4 (ErbB4). The gene symbol, ErbB, is derived from the name of a viral oncogene to which these receptors are homologous: erythroblastic leukemia viral oncogene. Insufficient ErbB signaling in humans is associated with the development of neurodegenerative diseases, such as multiple sclerosis and Alzheimer's disease, while excessive ErbB signaling is associated with the development of a wide variety of types of solid tumor.

ETS Like-1 protein Elk-1 is a protein that in humans is encoded by the ELK1. Elk-1 functions as a transcription activator. It is classified as a ternary complex factor (TCF), a subclass of the ETS family, which is characterized by a common protein domain that regulates DNA binding to target sequences. Elk1 plays important roles in various contexts, including long-term memory formation, drug addiction, Alzheimer's disease, Down syndrome, breast cancer, and depression.

Mitogen-activated protein kinase 7 also known as MAP kinase 7 is an enzyme that in humans is encoded by the MAPK7 gene.
Trk receptors are a family of tyrosine kinases that regulates synaptic strength and plasticity in the mammalian nervous system. Trk receptors affect neuronal survival and differentiation through several signaling cascades. However, the activation of these receptors also has significant effects on functional properties of neurons.

Dual specificity mitogen-activated protein kinase kinase 5 is an enzyme that in humans is encoded by the MAP2K5 gene.
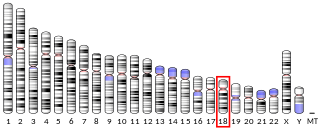
Mitogen-activated protein kinase 4 is an enzyme that in humans is encoded by the MAPK4 gene.
The Akt signaling pathway or PI3K-Akt signaling pathway is a signal transduction pathway that promotes survival and growth in response to extracellular signals. Key proteins involved are PI3K and Akt.

Tyrosine phosphorylation is the addition of a phosphate (PO43−) group to the amino acid tyrosine on a protein. It is one of the main types of protein phosphorylation. This transfer is made possible through enzymes called tyrosine kinases. Tyrosine phosphorylation is a key step in signal transduction and the regulation of enzymatic activity.
References
- ↑ Bohmer et al. "Cytoskeletal Integrity Is Required throughout the Mitogen Stimulation Phase of the Cell Cycle and Mediates the Anchorage-dependent Expression of Cyclin DI". January 1996, Molecular Biology of the Cell, Vol. 7, pp. 101-111.
- ↑ Foijer et al. "Mitogen requirement for cell cycle progression in the absence of pocket protein activity". December 2005, Cancer Cell, Vol. 8, pp. 455-466
- ↑ Gemberling et al. "Nrg1 is an injury-induced cardiomyocyte mitogen for the endogenous heart regeneration program in zebrafish". 1 April 2015, eLifeSciences.
- ↑ Leung et al. "Vascular Endothelial Growth Factor is a Secreted Angiogenic Mitogen". 8 December 1989, Science, Vol. 246, pp 1306-1309.
- 1 2 3 4 5 6 Morgan, David (2007). “The Cell Cycle: Principles of Control”. New Science Press.
- ↑ Waterfield, M., Scrace, G., Whittle, N. et al. Platelet-derived growth factor is structurally related to the putative transforming protein p28sis of simian sarcoma virus. Nature 304, 35–39 (1983) doi:10.1038/304035a0
- ↑ Mitri Z, Constantine T, O'Regan R (2012). "The HER2 Receptor in Breast Cancer: Pathophysiology, Clinical Use, and New Advances in Therapy". Chemotherapy Research and Practice. 2012: 743193
- ↑ Santen et al. "The role of mitogen-activated protein (MAP) kinase in breast cancer". February 2002, The Journal of Steroid Biochemistry and Molecular Biology, Vol. 80, pp. 239-256
- ↑ Barret, James (1980). Basic Immunology and its Medical Application (2 ed.). St.Louis: The C.V. Mosby Company. pp. 52–3. ISBN 978-0-8016-0495-9.
- ↑ Assenmacher, Mario; Avraham, Hava Karsenty; Avraham, Shalom; Bala, Shukal, eds. (2005), "Pokeweed Mitogen", Encyclopedic Reference of Immunotoxicology, Springer, p. 509, doi:10.1007/3-540-27806-0_1183, ISBN 978-3-540-27806-1
- ↑ Font-Nieves, M; Sans-Fons, MG (2012). "Induction of COX-2 enzyme and down-regulation of COX-1 expression by lipopolysaccharide (LPS) control prostaglandin E2 production in astrocytes". Journal of Biological Chemistry. 287 (9): 6454–68. doi: 10.1074/jbc.M111.327874 . PMC 3307308 . PMID 22219191.
- ↑ Casciani, V; Marinoni, E (2008). "Opposite effect of phorbol ester PMA on PTGS2 and PGDH mRNA expression in human chorion trophoblast cells". Reproductive Sciences. 15 (1): 40–50. doi:10.1177/1933719107309647. PMID 18212353. S2CID 10706385.