Related Research Articles
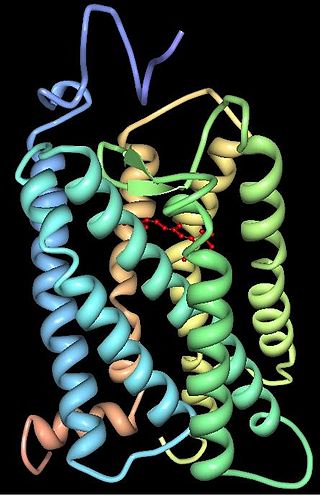
Rhodopsin, also known as visual purple, is a protein encoded by the RHO gene and a G-protein-coupled receptor (GPCR). It is the opsin of the rod cells in the retina and a light-sensitive receptor protein that triggers visual phototransduction in rods. Rhodopsin mediates dim light vision and thus is extremely sensitive to light. When rhodopsin is exposed to light, it immediately photobleaches. In humans, it is regenerated fully in about 30 minutes, after which the rods are more sensitive. Defects in the rhodopsin gene cause eye diseases such as retinitis pigmentosa and congenital stationary night blindness.

A photoreceptor cell is a specialized type of neuroepithelial cell found in the retina that is capable of visual phototransduction. The great biological importance of photoreceptors is that they convert light into signals that can stimulate biological processes. To be more specific, photoreceptor proteins in the cell absorb photons, triggering a change in the cell's membrane potential.
Bacteriorhodopsin (Bop) is a protein used by Archaea, most notably by haloarchaea, a class of the Euryarchaeota. It acts as a proton pump; that is, it captures light energy and uses it to move protons across the membrane out of the cell. The resulting proton gradient is subsequently converted into chemical energy.

Rod cells are photoreceptor cells in the retina of the eye that can function in lower light better than the other type of visual photoreceptor, cone cells. Rods are usually found concentrated at the outer edges of the retina and are used in peripheral vision. On average, there are approximately 92 million rod cells in the human retina. Rod cells are more sensitive than cone cells and are almost entirely responsible for night vision. However, rods have little role in color vision, which is the main reason why colors are much less apparent in dim light.
In visual physiology, adaptation is the ability of the retina of the eye to adjust to various levels of light. Natural night vision, or scotopic vision, is the ability to see under low-light conditions. In humans, rod cells are exclusively responsible for night vision as cone cells are only able to function at higher illumination levels. Night vision is of lower quality than day vision because it is limited in resolution and colors cannot be discerned; only shades of gray are seen. In order for humans to transition from day to night vision they must undergo a dark adaptation period of up to two hours in which each eye adjusts from a high to a low luminescence "setting", increasing sensitivity hugely, by many orders of magnitude. This adaptation period is different between rod and cone cells and results from the regeneration of photopigments to increase retinal sensitivity. Light adaptation, in contrast, works very quickly, within seconds.

Guanylate cyclase is a lyase enzyme that converts guanosine triphosphate (GTP) to cyclic guanosine monophosphate (cGMP) and pyrophosphate:
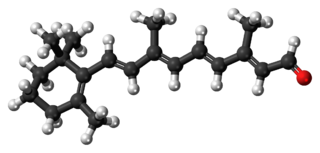
Retinal is a polyene chromophore. Retinal, bound to proteins called opsins, is the chemical basis of visual phototransduction, the light-detection stage of visual perception (vision).

Melanopsin is a type of photopigment belonging to a larger family of light-sensitive retinal proteins called opsins and encoded by the gene Opn4. In the mammalian retina, there are two additional categories of opsins, both involved in the formation of visual images: rhodopsin and photopsin in the rod and cone photoreceptor cells, respectively.
Photopigments are unstable pigments that undergo a chemical change when they absorb light. The term is generally applied to the non-protein chromophore moiety of photosensitive chromoproteins, such as the pigments involved in photosynthesis and photoreception. In medical terminology, "photopigment" commonly refers to the photoreceptor proteins of the retina.
Animal opsins are G-protein-coupled receptors and a group of proteins made light-sensitive via a chromophore, typically retinal. When bound to retinal, opsins become Retinylidene proteins, but are usually still called opsins regardless. Most prominently, they are found in photoreceptor cells of the retina. Five classical groups of opsins are involved in vision, mediating the conversion of a photon of light into an electrochemical signal, the first step in the visual transduction cascade. Another opsin found in the mammalian retina, melanopsin, is involved in circadian rhythms and pupillary reflex but not in vision. Humans have in total nine opsins. Beside vision and light perception, opsins may also sense temperature, sound, or chemicals.
Visual phototransduction is the sensory transduction process of the visual system by which light is detected to yield nerve impulses in the rod cells and cone cells in the retina of the eye in humans and other vertebrates. It relies on the visual cycle, a sequence of biochemical reactions in which a molecule of retinal bound to opsin undergoes photoisomerization, initiates a cascade that signals detection of the photon, and is indirectly restored to its photosensitive isomer for reuse. Phototransduction in some invertebrates such as fruit flies relies on similar processes.
Photoreceptor proteins are light-sensitive proteins involved in the sensing and response to light in a variety of organisms. Some examples are rhodopsin in the photoreceptor cells of the vertebrate retina, phytochrome in plants, and bacteriorhodopsin and bacteriophytochromes in some bacteria. They mediate light responses as varied as visual perception, phototropism and phototaxis, as well as responses to light-dark cycles such as circadian rhythm and other photoperiodisms including control of flowering times in plants and mating seasons in animals.

The eyespot apparatus is a photoreceptive organelle found in the flagellate or (motile) cells of green algae and other unicellular photosynthetic organisms such as euglenids. It allows the cells to sense light direction and intensity and respond to it, prompting the organism to either swim towards the light, or away from it. A related response occurs when cells are briefly exposed to high light intensity, causing the cell to stop, briefly swim backwards, then change swimming direction. Eyespot-mediated light perception helps the cells in finding an environment with optimal light conditions for photosynthesis. Eyespots are the simplest and most common "eyes" found in nature, composed of photoreceptors and areas of bright orange-red red pigment granules. Signals relayed from the eyespot photoreceptors result in alteration of the beating pattern of the flagella, generating a phototactic response.
The visual cycle is a process in the retina that replenishes the molecule retinal for its use in vision. Retinal is the chromophore of most visual opsins, meaning it captures the photons to begin the phototransduction cascade. When the photon is absorbed, the 11-cis retinal photoisomerizes into all-trans retinal as it is ejected from the opsin protein. Each molecule of retinal must travel from the photoreceptor cell to the RPE and back in order to be refreshed and combined with another opsin. This closed enzymatic pathway of 11-cis retinal is sometimes called Wald's visual cycle after George Wald (1906–1997), who received the Nobel Prize in 1967 for his work towards its discovery.

Peropsin, a visual pigment-like receptor, is a protein that in humans is encoded by the RRH gene. It belongs like other animal opsins to the G protein-coupled receptors. Even so, the first peropsins were already discovered in mice and humans in 1997, not much is known about them.
RPE-retinal G protein-coupled receptor also known as RGR-opsin is a protein that in humans is encoded by the RGR gene. RGR-opsin is a member of the rhodopsin-like receptor subfamily of GPCR. Like other opsins which bind retinaldehyde, it contains a conserved lysine residue in the seventh transmembrane domain. RGR-opsin comes in different isoforms produced by alternative splicing.
G-protein-coupled receptor kinase 7 is a serine/threonine-specific protein kinase involved in phototransduction. This enzyme catalyses the phosphorylation of cone (color) photopsins in retinal cones during high acuity color vision primarily in the fovea.

Microbial rhodopsins, also known as bacterial rhodopsins are retinal-binding proteins that provide light-dependent ion transport and sensory functions in halophilic and other bacteria. They are integral membrane proteins with seven transmembrane helices, the last of which contains the attachment point for retinal.

Georg Nagel is a biophysicist and professor at the Department for Neurophysiology at the University of Würzburg in Germany. His research is focused on microbial photoreceptors and the development of optogenetic tools.
Vertebrate visual opsins are a subclass of ciliary opsins and mediate vision in vertebrates. They include the opsins in human rod and cone cells. They are often abbreviated to opsin, as they were the first opsins discovered and are still the most widely studied opsins.
References
- 1 2 Spudich, John L.; Yang, Chii-Shen; Jung, Kwang-Hwan; Spudich, Elena N. (November 2000). "Retinylidene Proteins: Structures and Functions from Archaea to Humans". Annual Review of Cell and Developmental Biology. 16 (1): 365–392. doi:10.1146/annurev.cellbio.16.1.365. PMID 11031241.
- ↑ Mason, Peggy (26 May 2011). Medical Neurobiology. OUP USA. p. 375. ISBN 978-0-19-533997-0 . Retrieved 21 September 2015.
- ↑ Hara, Toshiaki J.; Zielinski, Barbara (17 October 2006). Fish Physiology: Sensory Systems Neuroscience: Sensory Systems Neuroscience. Academic Press. p. 183. ISBN 978-0-08-046961-4 . Retrieved 21 September 2015.
- ↑ Tsukamoto, T.; Inoue, K.; Kandori, H.; Sudo, Y. (2013). "Thermal and Spectroscopic Characterization of a Proton Pumping Rhodopsin from an Extreme Thermophile". Journal of Biological Chemistry. 288 (30): 21581–21592. doi: 10.1074/jbc.M113.479394 . ISSN 0021-9258. PMC 3724618 . PMID 23740255.
- ↑ Sakmar T (2002). "Structure of rhodopsin and the superfamily of seven-helical receptors: the same and not the same". Curr Opin Cell Biol. 14 (2): 189–95. doi:10.1016/S0955-0674(02)00306-X. PMID 11891118.
- 1 2 Bryant DA, Frigaard NU (November 2006). "Prokaryotic photosynthesis and phototrophy illuminated". Trends in Microbiology. 14 (11): 488–496. doi:10.1016/j.tim.2006.09.001. PMID 16997562.
- ↑ Nagel G, Szellas T, Kateriya S, Adeishvili N, Hegemann P, Bamberg E (2005). "Channelrhodopsins: directly light-gated cation channels". Biochem Soc Trans. 33 (Pt 4): 863–6. doi:10.1042/BST0330863. PMID 16042615.
- ↑ Gao S, Nagpal J, Schneider MW, Kozjak-Pavlovic V, Nagel G, Gottschalk A (September 2015). "Optogenetic manipulation of cGMP in cells and animals by the tightly light-regulated guanylyl-cyclase opsin CyclOp". Nature Communications. 6 (8046): 8046. Bibcode:2015NatCo...6.8046G. doi:10.1038/ncomms9046. PMC 4569695 . PMID 26345128.
- ↑ Scheib U, Stehfest K, Gee CE, Körschen HG, Fudim R, Oertner TG, Hegemann P (August 2015). "The rhodopsin-guanylyl cyclase of the aquatic fungus Blastocladiella emersonii enables fast optical control of cGMP signaling". Science Signaling. 8 (389): rs8. doi:10.1126/scisignal.aab0611. PMID 26268609. S2CID 13140205.
- ↑ Scheib U, Broser M, Constantin OM, Yang S, Gao S, Mukherjee S, et al. (May 2018). "Rhodopsin-cyclases for photocontrol of cGMP/cAMP and 2.3 Å structure of the adenylyl cyclase domain". Nature Communications. 9 (1): 2046. Bibcode:2018NatCo...9.2046S. doi:10.1038/s41467-018-04428-w. PMC 5967339 . PMID 29799525.
- ↑ "Superfamily: Bacterial photosystem II reaction centre, L and M subunits". SCOP.
- ↑ G Protein-Coupled Receptor Data Base
- 1 2 3 Terakita A (2005). "The opsins". Genome Biol. 6 (3): 213. doi:10.1186/gb-2005-6-3-213. PMC 1088937 . PMID 15774036.