Related Research Articles
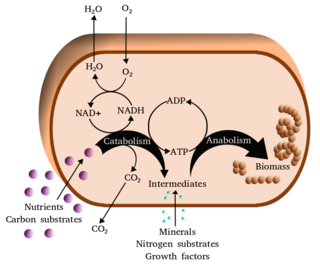
Metabolism is the set of life-sustaining chemical reactions in organisms. The three main functions of metabolism are: the conversion of the energy in food to energy available to run cellular processes; the conversion of food to building blocks of proteins, lipids, nucleic acids, and some carbohydrates; and the elimination of metabolic wastes. These enzyme-catalyzed reactions allow organisms to grow and reproduce, maintain their structures, and respond to their environments. The word metabolism can also refer to the sum of all chemical reactions that occur in living organisms, including digestion and the transportation of substances into and between different cells, in which case the above described set of reactions within the cells is called intermediary metabolism.

In biochemistry, a metabolic pathway is a linked series of chemical reactions occurring within a cell. The reactants, products, and intermediates of an enzymatic reaction are known as metabolites, which are modified by a sequence of chemical reactions catalyzed by enzymes. In most cases of a metabolic pathway, the product of one enzyme acts as the substrate for the next. However, side products are considered waste and removed from the cell.

A heterotroph is an organism that cannot produce its own food, instead taking nutrition from other sources of organic carbon, mainly plant or animal matter. In the food chain, heterotrophs are primary, secondary and tertiary consumers, but not producers. Living organisms that are heterotrophic include all animals and fungi, some bacteria and protists, and many parasitic plants. The term heterotroph arose in microbiology in 1946 as part of a classification of microorganisms based on their type of nutrition. The term is now used in many fields, such as ecology, in describing the food chain.

Acetyl-CoA is a molecule that participates in many biochemical reactions in protein, carbohydrate and lipid metabolism. Its main function is to deliver the acetyl group to the citric acid cycle to be oxidized for energy production.

Lignin is a class of complex organic polymers that form key structural materials in the support tissues of most plants. Lignins are particularly important in the formation of cell walls, especially in wood and bark, because they lend rigidity and do not rot easily. Chemically, lignins are polymers made by cross-linking phenolic precursors.
A biomolecule or biological molecule is loosely defined as a molecule produced by a living organism and essential to one or more typically biological processes. Biomolecules include large macromolecules such as proteins, carbohydrates, lipids, and nucleic acids, as well as small molecules such as vitamins and hormones. A more general name for this class of material is biological materials. Biomolecules are an important element of living organisms, those biomolecules are often endogenous, produced within the organism but organisms usually need exogenous biomolecules, for example certain nutrients, to survive.

Plant nutrition is the study of the chemical elements and compounds necessary for plant growth and reproduction, plant metabolism and their external supply. In its absence the plant is unable to complete a normal life cycle, or that the element is part of some essential plant constituent or metabolite. This is in accordance with Justus von Liebig's law of the minimum. The total essential plant nutrients include seventeen different elements: carbon, oxygen and hydrogen which are absorbed from the air, whereas other nutrients including nitrogen are typically obtained from the soil.

The metabolome refers to the complete set of small-molecule chemicals found within a biological sample. The biological sample can be a cell, a cellular organelle, an organ, a tissue, a tissue extract, a biofluid or an entire organism. The small molecule chemicals found in a given metabolome may include both endogenous metabolites that are naturally produced by an organism as well as exogenous chemicals that are not naturally produced by an organism.
Fatty acid metabolism consists of various metabolic processes involving or closely related to fatty acids, a family of molecules classified within the lipid macronutrient category. These processes can mainly be divided into (1) catabolic processes that generate energy and (2) anabolic processes where they serve as building blocks for other compounds.
Industrial fermentation is the intentional use of fermentation in manufacturing processes. In addition to the mass production of fermented foods and drinks, industrial fermentation has widespread applications in chemical industry. Commodity chemicals, such as acetic acid, citric acid, and ethanol are made by fermentation. Moreover, nearly all commercially produced industrial enzymes, such as lipase, invertase and rennet, are made by fermentation with genetically modified microbes. In some cases, production of biomass itself is the objective, as is the case for single-cell proteins, baker's yeast, and starter cultures for lactic acid bacteria used in cheesemaking.
The photosynthetic efficiency is the fraction of light energy converted into chemical energy during photosynthesis in green plants and algae. Photosynthesis can be described by the simplified chemical reaction

The microbial loop describes a trophic pathway where, in aquatic systems, dissolved organic carbon (DOC) is returned to higher trophic levels via its incorporation into bacterial biomass, and then coupled with the classic food chain formed by phytoplankton-zooplankton-nekton. In soil systems, the microbial loop refers to soil carbon. The term microbial loop was coined by Farooq Azam, Tom Fenchel et al. in 1983 to include the role played by bacteria in the carbon and nutrient cycles of the marine environment.
A xerophyte is a species of plant that has adaptations to survive in an environment with little liquid water. Examples of xerophytes include cacti, pineapple and some gymnosperm plants. The morphology and physiology of xerophytes are adapted to conserve water during dry periods. Some species called resurrection plants can survive long periods of extreme dryness or desiccation of their tissues, during which their metabolic activity may effectively shut down. Plants with such morphological and physiological adaptations are said to be xeromorphic. Xerophytes such as cacti are capable of withstanding extended periods of dry conditions as they have deep-spreading roots and capacity to store water. Their waxy, thorny leaves prevent loss of moisture.

Cellulose fibers are fibers made with ethers or esters of cellulose, which can be obtained from the bark, wood or leaves of plants, or from other plant-based material. In addition to cellulose, the fibers may also contain hemicellulose and lignin, with different percentages of these components altering the mechanical properties of the fibers.
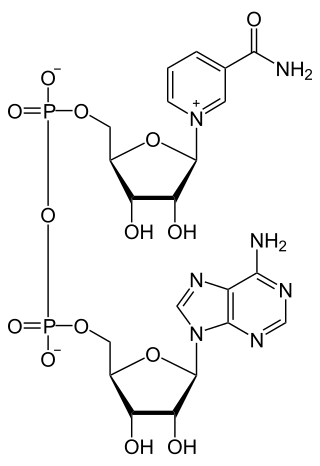
Cofactor engineering, a subset of metabolic engineering, is defined as the manipulation of the use of cofactors in an organism’s metabolic pathways. In cofactor engineering, the concentrations of cofactors are changed in order to maximize or minimize metabolic fluxes. This type of engineering can be used to optimize the production of a metabolite product or to increase the efficiency of a metabolic network. The use of engineering single celled organisms to create lucrative chemicals from cheap raw materials is growing, and cofactor engineering can play a crucial role in maximizing production. The field has gained more popularity in the past decade and has several practical applications in chemical manufacturing, bioengineering and pharmaceutical industries.
Biomass partitioning is the process by which plants divide their energy among their leaves, stems, roots, and reproductive parts. These four main components of the plant have important morphological roles: leaves take in CO2 and energy from the sun to create carbon compounds, stems grow above competitors to reach sunlight, roots absorb water and mineral nutrients from the soil while anchoring the plant, and reproductive parts facilitate the continuation of species. Plants partition biomass in response to limits or excesses in resources like sunlight, carbon dioxide, mineral nutrients, and water and growth is regulated by a constant balance between the partitioning of biomass between plant parts. An equilibrium between root and shoot growth occurs because roots need carbon compounds from photosynthesis in the shoot and shoots need nitrogen absorbed from the soil by roots. Allocation of biomass is put towards the limit to growth; a limit below ground will focus biomass to the roots and a limit above ground will favor more growth in the shoot.
Leaf expansion is a process by which plants make efficient use of the space around them by causing their leaves to enlarge, or wither. This process enables a plant to maximize its own biomass, whether it be due to increased surface area; which enables more sunlight to be absorbed by chloroplasts, driving the rate of photosynthesis upward, or it enables more stomata to be created on the leaf surface, allowing the plant to increase its carbon dioxide intake.
Biomass allocation is a concept in plant biology which indicates the relative proportion of plant biomass present in the different organs of a plant. It can also be used for whole plant communities.
Plant growth analysis refers to a set of concepts and equations by which changes in size of plants over time can be summarised and dissected in component variables. It is often applied in the analysis of growth of individual plants, but can also be used in a situation where crop growth is followed over time.

Acinetobacter baylyi is a bacterial species of the genus Acinetobacter. The species naming designation was given after the discovery of strains in activated sludge in Victoria, Australia, in 2003. A. baylyi is named after the late Dr. Ronald Bayly, an Australian microbiologist who contributed significantly to research on aromatic compound catabolism in diverse bacteria, including strains of Pseudomonas, Alcaligenes, and Acinetobacter. The new species designation in 2003 was found to apply to an already well-studied Acinetobacter strain known as ADP1, a derivative of a soil isolate characterized in 1969. Strain ADP1 was previously designated Acinetobacter sp. and Acinetobacter calcoaceticus. Research, particularly in the field of genetics, has established A. baylyi as a model organism.
References
- ↑ Tamiya, H (1932). "Zur Theorie des respiratorischen Quotienten nebst einer Bemerkung über den Einfluß der oxydoreduktiven Zellvorgänge auf den Gaswechsel der Zellen". Acta Phytochimica. 6: 227.
- ↑ Penning de Vries, Frits W.T.; Brunsting, A.H.M.; Van Laar, H.H. (June 1974). "Products, requirements and efficiency of biosynthesis a quantitative approach". Journal of Theoretical Biology. 45 (2): 339–377. doi:10.1016/0022-5193(74)90119-2. PMID 4367755.
- ↑ Williams, Kimberlyn; Field, Christopher B.; Mooney, Harold A. (February 1989). "Relationships Among Leaf Construction Cost, Leaf Longevity, and Light Environment in Rain-Forest Plants of the Genus Piper". The American Naturalist. 133 (2): 198–211. doi:10.1086/284910. S2CID 83629932.
- ↑ Poorter, Hendrik (1994). "Construction costs and payback time of biomass: A whole plant perspective". In Roy, Jacques; Garnier, Eric (eds.). A whole-plant perspective on Carbon-Nitrogen interactions. SPB Academic Publishing. pp. 111–127. ISBN 978-9051030860.
- ↑ Vertregt, N.; Penning de Vries, F.W.T. (September 1987). "A rapid method for determining the efficiency of biosynthesis of plant biomass". Journal of Theoretical Biology. 128 (1): 109–119. doi:10.1016/S0022-5193(87)80034-6.
- ↑ Griffin, K. L. (1994). "Calorimetric Estimates of Construction Cost and Their use in Ecological Studies". Functional Ecology. 8 (5): 551–562. doi:10.2307/2389915. JSTOR 2389915.
- ↑ Lafitte, H.R.; Loomis, R.S. (1988). "Calculation of Growth Yield, Growth Respiration and Heat Content of Grain Sorghum from Elemental and Proximal Analyses". Annals of Botany. 62 (4): 353–361. doi:10.1093/oxfordjournals.aob.a087667.
- ↑ Poorter, Hendrik; Villar, Rafael (1997). "The fate of acquired C in plants". In Bazzaz, Fakhri; Grace, John (eds.). Plant Resource Allocation. San Diego: Academic Press. pp. 39–72. ISBN 9780120834907.
- 1 2 Poorter, Hendrik; Pepin, Steeve; Rijkers, Toon; de Jong, Yvonne; Evans, John R.; Körner, Christian (2006). "Construction costs, chemical composition and payback time of high- and low-irradiance leaves". Journal of Experimental Botany. 57 (2): 355–371. doi: 10.1093/jxb/erj002 . PMID 16303828.
- ↑ Poorter, H.; Van Berkel, Y.; Baxter, R.; Den Hertog, J.; Dijkstra, P.; Gifford, R. M.; Griffin, K. L.; Roumet, C.; Roy, J.; Wong, S. C. (1997). "The effect of elevated CO2 on the chemical composition and construction costs of leaves of 27 C3 species". Plant, Cell and Environment. 20 (4): 472–482. doi:10.1046/j.1365-3040.1997.d01-84.x. hdl: 1874/13536 .
- ↑ Villar, Rafael; Merino, José (2001). "Comparison of leaf construction costs in woody species with differing leaf life-spans in contrasting ecosystems". New Phytologist. 151 (1): 213–226. doi: 10.1046/j.1469-8137.2001.00147.x . PMID 33873381.
- ↑ Poorter, H.; Bergkotte, M. (1992). "Chemical composition of 24 wild species differing in relative growth rate". Plant, Cell and Environment. 15 (2): 221–229. doi:10.1111/j.1365-3040.1992.tb01476.x.