Related Research Articles

In biology, a mutation is an alteration in the nucleic acid sequence of the genome of an organism, virus, or extrachromosomal DNA. Viral genomes contain either DNA or RNA. Mutations result from errors during DNA or viral replication, mitosis, or meiosis or other types of damage to DNA, which then may undergo error-prone repair, cause an error during other forms of repair, or cause an error during replication. Mutations may also result from insertion or deletion of segments of DNA due to mobile genetic elements.

Molecular evolution is the process of change in the sequence composition of cellular molecules such as DNA, RNA, and proteins across generations. The field of molecular evolution uses principles of evolutionary biology and population genetics to explain patterns in these changes. Major topics in molecular evolution concern the rates and impacts of single nucleotide changes, neutral evolution vs. natural selection, origins of new genes, the genetic nature of complex traits, the genetic basis of speciation, the evolution of development, and ways that evolutionary forces influence genomic and phenotypic changes.

Pseudogenes are nonfunctional segments of DNA that resemble functional genes. Most arise as superfluous copies of functional genes, either directly by gene duplication or indirectly by reverse transcription of an mRNA transcript. Pseudogenes are usually identified when genome sequence analysis finds gene-like sequences that lack regulatory sequences needed for transcription or translation, or whose coding sequences are obviously defective due to frameshifts or premature stop codons. Pseudogenes are a type of junk DNA.
Gene duplication is a major mechanism through which new genetic material is generated during molecular evolution. It can be defined as any duplication of a region of DNA that contains a gene. Gene duplications can arise as products of several types of errors in DNA replication and repair machinery as well as through fortuitous capture by selfish genetic elements. Common sources of gene duplications include ectopic recombination, retrotransposition event, aneuploidy, polyploidy, and replication slippage.

A gene family is a set of several similar genes, formed by duplication of a single original gene, and generally with similar biochemical functions. One such family are the genes for human hemoglobin subunits; the ten genes are in two clusters on different chromosomes, called the α-globin and β-globin loci. These two gene clusters are thought to have arisen as a result of a precursor gene being duplicated approximately 500 million years ago.
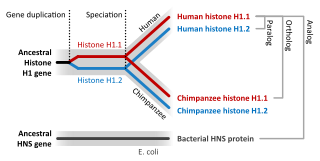
Sequence homology is the biological homology between DNA, RNA, or protein sequences, defined in terms of shared ancestry in the evolutionary history of life. Two segments of DNA can have shared ancestry because of three phenomena: either a speciation event (orthologs), or a duplication event (paralogs), or else a horizontal gene transfer event (xenologs).
In biology, the word gene can have several different meanings. The Mendelian gene is a basic unit of heredity and the molecular gene is a sequence of nucleotides in DNA that is transcribed to produce a functional RNA. There are two types of molecular genes: protein-coding genes and noncoding genes.

Directed evolution (DE) is a method used in protein engineering that mimics the process of natural selection to steer proteins or nucleic acids toward a user-defined goal. It consists of subjecting a gene to iterative rounds of mutagenesis, selection and amplification. It can be performed in vivo, or in vitro. Directed evolution is used both for protein engineering as an alternative to rationally designing modified proteins, as well as for experimental evolution studies of fundamental evolutionary principles in a controlled, laboratory environment.
Neutral mutations are changes in DNA sequence that are neither beneficial nor detrimental to the ability of an organism to survive and reproduce. In population genetics, mutations in which natural selection does not affect the spread of the mutation in a species are termed neutral mutations. Neutral mutations that are inheritable and not linked to any genes under selection will be lost or will replace all other alleles of the gene. That loss or fixation of the gene proceeds based on random sampling known as genetic drift. A neutral mutation that is in linkage disequilibrium with other alleles that are under selection may proceed to loss or fixation via genetic hitchhiking and/or background selection.

A gene family is a set of homologous genes within one organism. A gene cluster is a group of two or more genes found within an organism's DNA that encode similar polypeptides, or proteins, which collectively share a generalized function and are often located within a few thousand base pairs of each other. The size of gene clusters can vary significantly, from a few genes to several hundred genes. Portions of the DNA sequence of each gene within a gene cluster are found to be identical; however, the resulting protein of each gene is distinctive from the resulting protein of another gene within the cluster. Genes found in a gene cluster may be observed near one another on the same chromosome or on different, but homologous chromosomes. An example of a gene cluster is the Hox gene, which is made up of eight genes and is part of the Homeobox gene family.

Gene redundancy is the existence of multiple genes in the genome of an organism that perform the same function. Gene redundancy can result from gene duplication. Such duplication events are responsible for many sets of paralogous genes. When an individual gene in such a set is disrupted by mutation or targeted knockout, there can be little effect on phenotype as a result of gene redundancy, whereas the effect is large for the knockout of a gene with only one copy. Gene knockout is a method utilized in some studies aiming to characterize the maintenance and fitness effects functional overlap.
Evolutionary biology, in particular the understanding of how organisms evolve through natural selection, is an area of science with many practical applications. Creationists often claim that the theory of evolution lacks any practical applications; however, this claim has been refuted by scientists.
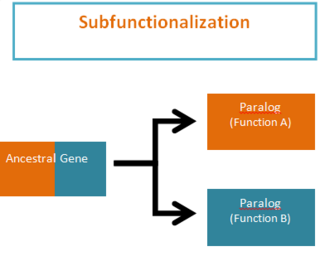
Subfunctionalization was proposed by Stoltzfus (1999) and Force et al. (1999) as one of the possible outcomes of functional divergence that occurs after a gene duplication event, in which pairs of genes that originate from duplication, or paralogs, take on separate functions. Subfunctionalization is a neutral mutation process of constructive neutral evolution; meaning that no new adaptations are formed. During the process of gene duplication paralogs simply undergo a division of labor by retaining different parts (subfunctions) of their original ancestral function. This partitioning event occurs because of segmental gene silencing leading to the formation of paralogs that are no longer duplicates, because each gene only retains a single function. It is important to note that the ancestral gene was capable of performing both functions and the descendant duplicate genes can now only perform one of the original ancestral functions.
A conserved non-coding sequence (CNS) is a DNA sequence of noncoding DNA that is evolutionarily conserved. These sequences are of interest for their potential to regulate gene production.
Bovine seminal RNase (BS-RNase) is a member of the ribonuclease superfamily produced by the bovine seminal vesicles. This enzyme can not be differentiated from its members distinctly since there are more features that this enzyme shares with its family members than features that it possess alone. The research on the question of how new functions arrive in proteins in evolution led the scientists to find an uncommon consequence for a usual biological event called gene conversion in the case of the ribonuclease (RNase) protein family. The most well-known member of this family, RNase A, is expressed in the pancreas of oxen. It serves to digest RNA in intestine, and evolved from bacteria fermenting in the stomach of the first ox. The homologous RNase, called seminal RNase, differs from RNase A by 23 amino acids and is expressed in seminal plasma in the concentration of 1-1.5 mg/ml, which constitutes more than 3% of the fluid protein content. Bovine seminal ribonuclease (BS-RNase) is a homologue of RNase A with specific antitumor activity.

Neofunctionalization, one of the possible outcomes of functional divergence, occurs when one gene copy, or paralog, takes on a totally new function after a gene duplication event. Neofunctionalization is an adaptive mutation process; meaning one of the gene copies must mutate to develop a function that was not present in the ancestral gene. In other words, one of the duplicates retains its original function, while the other accumulates molecular changes such that, in time, it can perform a different task.
Enzyme promiscuity is the ability of an enzyme to catalyse a fortuitous side reaction in addition to its main reaction. Although enzymes are remarkably specific catalysts, they can often perform side reactions in addition to their main, native catalytic activity. These promiscuous activities are usually slow relative to the main activity and are under neutral selection. Despite ordinarily being physiologically irrelevant, under new selective pressures these activities may confer a fitness benefit therefore prompting the evolution of the formerly promiscuous activity to become the new main activity. An example of this is the atrazine chlorohydrolase from Pseudomonas sp. ADP that evolved from melamine deaminase, which has very small promiscuous activity toward atrazine, a man-made chemical.

Epistasis is a phenomenon in genetics in which the effect of a gene mutation is dependent on the presence or absence of mutations in one or more other genes, respectively termed modifier genes. In other words, the effect of the mutation is dependent on the genetic background in which it appears. Epistatic mutations therefore have different effects on their own than when they occur together. Originally, the term epistasis specifically meant that the effect of a gene variant is masked by that of a different gene.
Constructive neutral evolution(CNE) is a theory that seeks to explain how complex systems can evolve through neutral transitions and spread through a population by chance fixation (genetic drift). Constructive neutral evolution is a competitor for both adaptationist explanations for the emergence of complex traits and hypotheses positing that a complex trait emerged as a response to a deleterious development in an organism. Constructive neutral evolution often leads to irreversible or "irremediable" complexity and produces systems which, instead of being finely adapted for performing a task, represent an excess complexity that has been described with terms such as "runaway bureaucracy" or even a "Rube Goldberg machine".
This glossary of genetics is a list of definitions of terms and concepts commonly used in the study of genetics and related disciplines in biology, including molecular biology, cell biology, and evolutionary biology. It is split across two articles:
References
- 1 2 Susumu Ohno (1970). Evolution by gene duplication. Springer-Verlag. ISBN 0-04-575015-7.
- 1 2 Andersson DI, Jerlström-Hultqvist J, Näsvall J. Evolution of new functions de novo and from preexisting genes. Cold Spring Harbor Perspectives in Biology. 2015 Jun 1;7(6):a017996.
- ↑ Lynch, M; et al. (2000). "The evolutionary fate and consequences of duplicate genes". Science. 290 (5494): 1151–2254. Bibcode:2000Sci...290.1151L. doi:10.1126/science.290.5494.1151. PMID 11073452.
- 1 2 3 Force, A.; et al. (1999). "Preservation of duplicate genes by complementary, degenerative mutations". Genetics. 151 (4): 1531–1545. doi:10.1093/genetics/151.4.1531. PMC 1460548 . PMID 10101175.
- 1 2 Bergthorsson U, Andersson DI, Roth JR (2007). "Ohno's dilemma: Evolution of new genes under continuous selection". PNAS. 104 (43): 17004–17009. Bibcode:2007PNAS..10417004B. doi: 10.1073/pnas.0707158104 . PMC 2040452 . PMID 17942681.
- ↑ Grauer, Dan; Li, Wen-Hsuing (2000). Fundamentals of molecular evolution . Sunderland, MA: Sinauer. pp. 282–283. ISBN 0-87893-266-6.
- ↑ Bergthorsson U, Andersson DI, Roth JR. Ohno's dilemma: evolution of new genes under continuous selection. Proceedings of the National Academy of Sciences. 2007 Oct 23;104(43):17004-9.
- ↑ Hittinger CT, Carroll SB (2007). "Gene duplication and the adaptive evolution of a classic genetic switch". Nature. 449 (7163): 677–81. Bibcode:2007Natur.449..677H. doi:10.1038/nature06151. PMID 17928853. S2CID 4418250.