
The cerebellum is a major feature of the hindbrain of all vertebrates. Although usually smaller than the cerebrum, in some animals such as the mormyrid fishes it may be as large as it or even larger. In humans, the cerebellum plays an important role in motor control. It may also be involved in some cognitive functions such as attention and language as well as emotional control such as regulating fear and pleasure responses, but its movement-related functions are the most solidly established. The human cerebellum does not initiate movement, but contributes to coordination, precision, and accurate timing: it receives input from sensory systems of the spinal cord and from other parts of the brain, and integrates these inputs to fine-tune motor activity. Cerebellar damage produces disorders in fine movement, equilibrium, posture, and motor learning in humans.

The inferior olivary nucleus (ION), is a structure found in the medulla oblongata underneath the superior olivary nucleus. In vertebrates, the ION is known to coordinate signals from the spinal cord to the cerebellum to regulate motor coordination and learning. These connections have been shown to be tightly associated, as degeneration of either the cerebellum or the ION results in degeneration of the other.
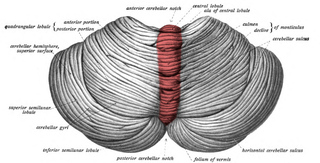
The cerebellar vermis is located in the medial, cortico-nuclear zone of the cerebellum, which is in the posterior fossa of the cranium. The primary fissure in the vermis curves ventrolaterally to the superior surface of the cerebellum, dividing it into anterior and posterior lobes. Functionally, the vermis is associated with bodily posture and locomotion. The vermis is included within the spinocerebellum and receives somatic sensory input from the head and proximal body parts via ascending spinal pathways.

Basket cells are inhibitory GABAergic interneurons of the brain, found throughout different regions of the cortex and cerebellum.
Climbing fibers are the name given to a series of neuronal projections from the inferior olivary nucleus located in the medulla oblongata.

Purkinje cells, or Purkinje neurons, are a class of GABAergic inhibitory neurons located in the cerebellum. They are named after their discoverer, Czech anatomist Jan Evangelista Purkyně, who characterized the cells in 1839.

There are four deep cerebellar nuclei embedded in the white matter of the medullary centre. The nuclei are the fastigial, globose, emboliform, and dentate nuclei.
The dentate nucleus is a cluster of neurons, or nerve cells, in the central nervous system that has a dentate – tooth-like or serrated – edge. It is located within the deep white matter of each cerebellar hemisphere, and it is the largest single structure linking the cerebellum to the rest of the brain. It is the largest and most lateral, or farthest from the midline, of the four pairs of deep cerebellar nuclei, the others being the globose and emboliform nuclei, which together are referred to as the interposed nucleus, and the fastigial nucleus. The dentate nucleus is responsible for the planning, initiation and control of voluntary movements. The dorsal region of the dentate nucleus contains output channels involved in motor function, which is the movement of skeletal muscle, while the ventral region contains output channels involved in nonmotor function, such as conscious thought and visuospatial function.

The globose nucleus is one of the deep cerebellar nuclei. It is located medial to the emboliform nucleus, and lateral to the fastigial nucleus. The globose nucleus and emboliform nucleus are known collectively as the interposed nuclei.
The fastigial nucleus is located in the cerebellum. It is one of the four deep cerebellar nuclei, and is grey matter embedded in the white matter of the cerebellum.
Dysmetria is a lack of coordination of movement typified by the undershoot or overshoot of intended position with the hand, arm, leg, or eye. It is a type of ataxia. It can also include an inability to judge distance or scale.

The flocculus is a small lobe of the cerebellum at the posterior border of the middle cerebellar peduncle anterior to the biventer lobule. Like other parts of the cerebellum, the flocculus is involved in motor control. It is an essential part of the vestibulo-ocular reflex, and aids in the learning of basic motor skills in the brain.
Cerebellar ataxia is a form of ataxia originating in the cerebellum. Non-progressive congenital ataxia (NPCA) is a classical presentation of cerebral ataxias.
Mossy fibers are one of the major inputs to cerebellum. There are many sources of this pathway, the largest of which is the cerebral cortex, which sends input to the cerebellum via the pontocerebellar pathway. Other contributors include the vestibular nerve and nuclei, the spinal cord, the reticular formation, and feedback from deep cerebellar nuclei. Axons enter the cerebellum via the middle and inferior cerebellar peduncles, where some branch to make contact with deep cerebellar nuclei. They ascend into the white matter of the cerebellum, where each axon branches to innervate granule cells in several cerebellar folia.
The cerebellothalamic tract or the tractus cerebellothalamicus, is part of the superior cerebellar peduncle. It originates in the cerebellar nuclei, crosses completely in the decussation of the superior cerebellar peduncle, bypasses the red nucleus, and terminates in posterior division of ventral lateral nucleus of thalamus. The ventrolateral nucleus has different divisions and distinct connections, mostly with frontal and parietal lobes. The primary motor cortex and premotor cortex get information from the ventrolateral nucleus projections originating in the interposed nucleus and dentate nuclei. Other dentate nucleus projections via thalamic pathway transmit information to prefrontal cortex and posterior parietal cortex. The cerebellum sends thalamocortical projections and in addition may also send connections from the thalamus to association areas serving cognitive and affective functions.
The anatomy of the cerebellum can be viewed at three levels. At the level of gross anatomy, the cerebellum consists of a tightly folded and crumpled layer of cortex, with white matter underneath, several deep nuclei embedded in the white matter, and a fluid-filled ventricle in the middle. At the intermediate level, the cerebellum and its auxiliary structures can be broken down into several hundred or thousand independently functioning modules or compartments known as microzones. At the microscopic level, each module consists of the same small set of neuronal elements, laid out with a highly stereotyped geometry.
The name granule cell has been used for a number of different types of neurons whose only common feature is that they all have very small cell bodies. Granule cells are found within the granular layer of the cerebellum, the dentate gyrus of the hippocampus, the superficial layer of the dorsal cochlear nucleus, the olfactory bulb, and the cerebral cortex.

The rhombic lip is a posterior section of the developing metencephalon which can be recognized transiently within the vertebrate embryo. It extends posteriorly from the roof of the fourth ventricle to dorsal neuroepithelial cells. The rhombic lip can be divided into eight structural units based on rhombomeres 1-8 (r1-r8), which can be recognized at early stages of hindbrain development. Producing granule cells and five brainstem nuclei, the rhombic lip plays an important role in developing a complex cerebellar neural system.
Cerebellar cognitive affective syndrome (CCAS), also called Schmahmann's syndrome is a condition that follows from lesions (damage) to the cerebellum of the brain. It refers to a constellation of deficits in the cognitive domains of executive function, spatial cognition, language, and affect resulting from damage to the cerebellum. Impairments of executive function include problems with planning, set-shifting, abstract reasoning, verbal fluency, and working memory, and there is often perseveration, distractibility and inattention. Language problems include dysprosodia, agrammatism and mild anomia. Deficits in spatial cognition produce visual–spatial disorganization and impaired visual–spatial memory. Personality changes manifest as blunting of affect or disinhibited and inappropriate behavior. These cognitive impairments result in an overall lowering of intellectual function. CCAS challenges the traditional view of the cerebellum being responsible solely for regulation of motor functions. It is now thought that the cerebellum is responsible for monitoring both motor and nonmotor functions. The nonmotor deficits described in CCAS are believed to be caused by dysfunction in cerebellar connections to the cerebral cortex and limbic system.

ATP/GTP binding protein 1 is gene that encodes the protein known as cytosolic carboxypeptidase 1 (CCP1), originally named NNA1. Mice with a naturally occurring mutation of the Agtpbp1 gene are known as pcd mice.