
The facial nerve, also known as the seventh cranial nerve, cranial nerve VII, or simply CN VII, is a cranial nerve that emerges from the pons of the brainstem, controls the muscles of facial expression, and functions in the conveyance of taste sensations from the anterior two-thirds of the tongue. The nerve typically travels from the pons through the facial canal in the temporal bone and exits the skull at the stylomastoid foramen. It arises from the brainstem from an area posterior to the cranial nerve VI and anterior to cranial nerve VIII.
Articles related to anatomy include:

The brainstem is the stalk-like part of the brain that interconnects the cerebrum and diencephalon with the spinal cord. In the human brain, the brainstem is composed of the midbrain, the pons, and the medulla oblongata. The midbrain is continuous with the thalamus of the diencephalon through the tentorial notch.

In neuroanatomy, the trigeminal nerve (lit. triplet nerve), also known as the fifth cranial nerve, cranial nerve V, or simply CN V, is a cranial nerve responsible for sensation in the face and motor functions such as biting and chewing; it is the most complex of the cranial nerves. Its name (trigeminal, from Latin tri- 'three', and -geminus 'twin') derives from each of the two nerves (one on each side of the pons) having three major branches: the ophthalmic nerve (V1), the maxillary nerve (V2), and the mandibular nerve (V3). The ophthalmic and maxillary nerves are purely sensory, whereas the mandibular nerve supplies motor as well as sensory (or "cutaneous") functions. Adding to the complexity of this nerve is that autonomic nerve fibers as well as special sensory fibers (taste) are contained within it.
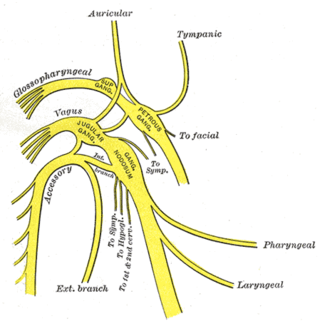
The glossopharyngeal nerve, also known as the ninth cranial nerve, cranial nerve IX, or simply CN IX, is a cranial nerve that exits the brainstem from the sides of the upper medulla, just anterior to the vagus nerve. Being a mixed nerve (sensorimotor), it carries afferent sensory and efferent motor information. The motor division of the glossopharyngeal nerve is derived from the basal plate of the embryonic medulla oblongata, whereas the sensory division originates from the cranial neural crest.

The hypoglossal nerve, also known as the twelfth cranial nerve, cranial nerve XII, or simply CN XII, is a cranial nerve that innervates all the extrinsic and intrinsic muscles of the tongue except for the palatoglossus, which is innervated by the vagus nerve. CN XII is a nerve with a sole motor function. The nerve arises from the hypoglossal nucleus in the medulla as a number of small rootlets, pass through the hypoglossal canal and down through the neck, and eventually passes up again over the tongue muscles it supplies into the tongue.

The internal capsule is a white matter structure situated in the inferomedial part of each cerebral hemisphere of the brain. It carries information past the basal ganglia, separating the caudate nucleus and the thalamus from the putamen and the globus pallidus. The internal capsule contains both ascending and descending axons, going to and coming from the cerebral cortex. It also separates the caudate nucleus and the putamen in the dorsal striatum, a brain region involved in motor and reward pathways.

The pyramidal tracts include both the corticobulbar tract and the corticospinal tract. These are aggregations of efferent nerve fibers from the upper motor neurons that travel from the cerebral cortex and terminate either in the brainstem (corticobulbar) or spinal cord (corticospinal) and are involved in the control of motor functions of the body.

The spinothalamic tract is a part of the anterolateral system or the ventrolateral system, a sensory pathway to the thalamus. From the ventral posterolateral nucleus in the thalamus, sensory information is relayed upward to the somatosensory cortex of the postcentral gyrus.
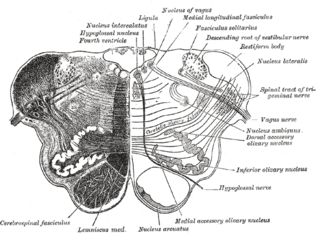
The nucleus ambiguus is a group of large motor neurons, situated deep in the medullary reticular formation named by Jacob Clarke. The nucleus ambiguus contains the cell bodies of neurons that innervate the muscles of the soft palate, pharynx, and larynx which are associated with speech and swallowing. As well as motor neurons, the nucleus ambiguus contains preganglionic parasympathetic neurons which innervate postganglionic parasympathetic neurons in the heart.

In neuroanatomy, the corticobulbartract is a two-neuron white matter motor pathway connecting the motor cortex in the cerebral cortex to the medullary pyramids, which are part of the brainstem's medulla oblongata region, and are primarily involved in carrying the motor function of the non-oculomotor cranial nerves. The corticobulbar tract is one of the pyramidal tracts, the other being the corticospinal tract.

The precentral gyrus is a prominent gyrus on the surface of the posterior frontal lobe of the brain. It is the site of the primary motor cortex that in humans is cytoarchitecturally defined as Brodmann area 4.

The dentate nucleus is a cluster of neurons, or nerve cells, in the central nervous system that has a dentate – tooth-like or serrated – edge. It is located within the deep white matter of each cerebellar hemisphere, and it is the largest single structure linking the cerebellum to the rest of the brain. It is the largest and most lateral, or farthest from the midline, of the four pairs of deep cerebellar nuclei, the others being the globose and emboliform nuclei, which together are referred to as the interposed nucleus, and the fastigial nucleus. The dentate nucleus is responsible for the planning, initiation and control of voluntary movements. The dorsal region of the dentate nucleus contains output channels involved in motor function, which is the movement of skeletal muscle, while the ventral region contains output channels involved in nonmotor function, such as conscious thought and visuospatial function.
The vestibulospinal tract is a neural tract in the central nervous system. Specifically, it is a component of the extrapyramidal system and is classified as a component of the medial pathway. Like other descending motor pathways, the vestibulospinal fibers of the tract relay information from nuclei to motor neurons. The vestibular nuclei receive information through the vestibulocochlear nerve about changes in the orientation of the head. The nuclei relay motor commands through the vestibulospinal tract. The function of these motor commands is to alter muscle tone, extend, and change the position of the limbs and head with the goal of supporting posture and maintaining balance of the body and head.

The cochlear nuclear (CN) complex comprises two cranial nerve nuclei in the human brainstem, the ventral cochlear nucleus (VCN) and the dorsal cochlear nucleus (DCN). The ventral cochlear nucleus is unlayered whereas the dorsal cochlear nucleus is layered. Auditory nerve fibers, fibers that travel through the auditory nerve carry information from the inner ear, the cochlea, on the same side of the head, to the nerve root in the ventral cochlear nucleus. At the nerve root the fibers branch to innervate the ventral cochlear nucleus and the deep layer of the dorsal cochlear nucleus. All acoustic information thus enters the brain through the cochlear nuclei, where the processing of acoustic information begins. The outputs from the cochlear nuclei are received in higher regions of the auditory brainstem.
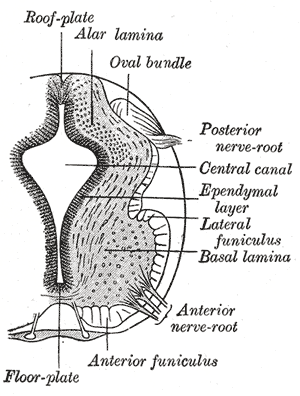
Alpha (α) motor neurons (also called alpha motoneurons), are large, multipolar lower motor neurons of the brainstem and spinal cord. They innervate extrafusal muscle fibers of skeletal muscle and are directly responsible for initiating their contraction. Alpha motor neurons are distinct from gamma motor neurons, which innervate intrafusal muscle fibers of muscle spindles.

Foix–Chavany–Marie Syndrome (FCMS), also known as bilateral opercular syndrome, is a neuropathological disorder characterized by paralysis of the facial, tongue, pharynx, and masticatory muscles of the mouth that aid in chewing. The disorder is primarily caused by thrombotic and embolic strokes, which cause a deficiency of oxygen in the brain. As a result, bilateral lesions may form in the junctions between the frontal lobe and temporal lobe, the parietal lobe and cortical lobe, or the subcortical region of the brain. FCMS may also arise from defects existing at birth that may be inherited or nonhereditary. Symptoms of FCMS can be present in a person of any age and it is diagnosed using automatic-voluntary dissociation assessment, psycholinguistic testing, neuropsychological testing, and brain scanning. Treatment for FCMS depends on the onset, as well as on the severity of symptoms, and it involves a multidisciplinary approach.
Central facial palsy is a symptom or finding characterized by paralysis or paresis of the lower half of one side of the face. It usually results from damage to upper motor neurons of the facial nerve.

In the ventral cochlear nucleus (VCN), auditory nerve fibers enter the brain via the nerve root in the VCN. The ventral cochlear nucleus is divided into the anterior ventral (anteroventral) cochlear nucleus (AVCN) and the posterior ventral (posteroventral) cochlear nucleus (PVCN). In the VCN, auditory nerve fibers bifurcate, the ascending branch innervates the AVCN and the descending branch innervates the PVCN and then continue to the dorsal cochlear nucleus. The orderly innervation by auditory nerve fibers gives the AVCN a tonotopic organization along the dorsoventral axis. Fibers that carry information from the apex of the cochlea that are tuned to low frequencies contact neurons in the ventral part of the AVCN; those that carry information from the base of the cochlea that are tuned to high frequencies contact neurons in the dorsal part of the AVCN. Several populations of neurons populate the AVCN. Bushy cells receive input from auditory nerve fibers through particularly large endings called end bulbs of Held. They contact stellate cells through more conventional boutons.
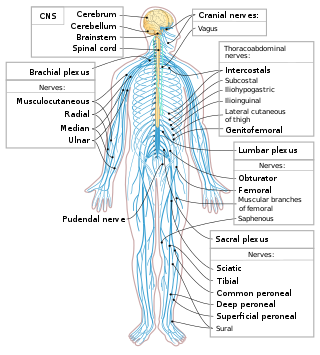
The spinal cord is a long, thin, tubular structure made up of nervous tissue that extends from the medulla oblongata in the brainstem to the lumbar region of the vertebral column (backbone) of vertebrate animals. The center of the spinal cord is hollow and contains a structure called central canal, which contains cerebrospinal fluid. The spinal cord is also covered by meninges and enclosed by the neural arches. Together, the brain and spinal cord make up the central nervous system (CNS).




