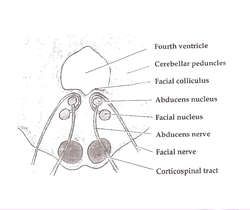
The pons is part of the brainstem that in humans and other mammals, lies inferior to the midbrain, superior to the medulla oblongata and anterior to the cerebellum.

The brainstem is the posterior stalk-like part of the brain that connects the cerebrum with the spinal cord. In the human brain the brainstem is composed of the midbrain, the pons, and the medulla oblongata. The midbrain is continuous with the thalamus of the diencephalon through the tentorial notch, and sometimes the diencephalon is included in the brainstem.
In neuroanatomy, a nucleus is a cluster of neurons in the central nervous system, located deep within the cerebral hemispheres and brainstem. The neurons in one nucleus usually have roughly similar connections and functions. Nuclei are connected to other nuclei by tracts, the bundles (fascicles) of axons extending from the cell bodies. A nucleus is one of the two most common forms of nerve cell organization, the other being layered structures such as the cerebral cortex or cerebellar cortex. In anatomical sections, a nucleus shows up as a region of gray matter, often bordered by white matter. The vertebrate brain contains hundreds of distinguishable nuclei, varying widely in shape and size. A nucleus may itself have a complex internal structure, with multiple types of neurons arranged in clumps (subnuclei) or layers.
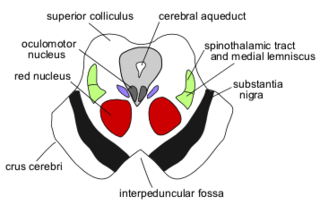
The midbrain or mesencephalon is the uppermost portion of the brainstem connecting the diencephalon and cerebrum with the pons. It consists of the cerebral peduncles, tegmentum, and tectum.
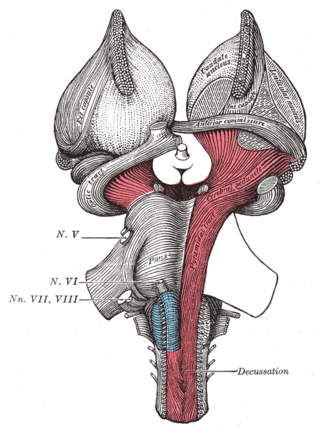
The cerebral peduncles are the two stalks that attach the cerebrum to the brainstem. They are structures at the front of the midbrain which arise from the ventral pons and contain the large ascending (sensory) and descending (motor) tracts that run to and from the cerebrum from the pons. Mainly, the three common areas that give rise to the cerebral peduncles are the cerebral cortex, the spinal cord and the cerebellum. The region includes the tegmentum, crus cerebri and pretectum. By this definition, the cerebral peduncles are also known as the basis pedunculi, while the large ventral bundle of efferent fibers is referred to as the cerebral crus or the pes pedunculi.

The internal capsule is a paired white matter structure, as a two-way tract, carrying ascending and descending fibers, to and from the cerebral cortex. The internal capsule is situated in the inferomedial part of each cerebral hemisphere of the brain. It carries information past the subcortical basal ganglia. As it courses it separates the caudate nucleus and the thalamus from the putamen and the globus pallidus. It also separates the caudate nucleus and the putamen in the dorsal striatum, a brain region involved in motor and reward pathways.

The reticular formation is a set of interconnected nuclei in the brainstem that spans from the lower end of the medulla oblongata to the upper end of the midbrain. The neurons of the reticular formation make up a complex set of neural networks in the core of the brainstem. The reticular formation is made up of a diffuse net-like formation of reticular nuclei which is not well-defined. It may be seen as being made up of all the interspersed cells in the brainstem between the more compact and named structures.

The pontine nuclei are all the neurons of the ventral pons. Corticopontine fibres project from the primary motor cortex to the ipsilateral pontine nucleus; pontocerebellar fibers then relay the information to the contralateral cerebellum via the middle cerebellar peduncle.
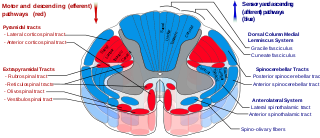
The spinocerebellar tracts are nerve tracts originating in the spinal cord and terminating in the same side (ipsilateral) of the cerebellum. The two main tracts are the dorsal spinocerebellar tract, and the ventral spinocerebellar tract. Both of these tracts are located in the peripheral region of the lateral funiculi. Other tracts are the rostral spinocerebellar tract, and the cuneocerebellar tract.

The dentate nucleus refer to a pair of deep cerebellar nuclei deep within the white matter of the cerebellum of the brain with a dentate – tooth-like or serrated – edge. The dentate forms the largest pathway between the cerebellum and the remainder of the brain. It is the largest and most lateral of the four pairs of deep cerebellar nuclei, the others being the globose and emboliform nuclei, which together are referred to as the interposed nucleus, and the fastigial nucleus.

The globose nucleus is one of the deep cerebellar nuclei. It is located medial to the emboliform nucleus, and lateral to the fastigial nucleus. The globose nucleus and emboliform nucleus are known collectively as the interposed nuclei.
The interposed nucleus is the combined paired globose and emboliform nuclei, on either side of the cerebellum. It is located in the roof of the fourth ventricle, lateral to the fastigial nucleus. The emboliform nucleus is the anterior interposed nucleus, and the globose nucleus is the posterior interposed nucleus.

The flocculus is a small lobe of the cerebellum at the posterior border of the middle cerebellar peduncle anterior to the biventer lobule. Like other parts of the cerebellum, the flocculus is involved in motor control. It is an essential part of the vestibulo-ocular reflex, and aids in the learning of basic motor skills in the brain.

In the human brain, the superior cerebellar peduncle is one of the three paired cerebellar peduncles of bundled fibers that connect the cerebellum to the brainstem. The superior cerebellar peduncle connects to the midbrain. It consists mainly of efferent fibers, the cerebellothalamic tract that runs from a cerebellar hemisphere to the contralateral thalamus, and the cerebellorubral tract that runs from a cerebellar hemisphere to the red nucleus. It also contains afferent tracts, most prominent of which is the ventral spinocerebellar tract. Other afferent tracts are the ventral trigeminal tract, tectocerebellar fibers, and noradrenergic fibers from the locus coeruleus. The superior peduncle emerges from the upper and medial parts of the white matter of each cerebellar hemisphere and is placed under cover of the upper part of the cerebellum.

The middle cerebellar peduncle is one of three paired cerebellar peduncles connecting the brainstem to the cerebellum. The connection is from the pons. It connects the pons to the cerebellum, with fibres originating from the pontine nuclei, and travelling to the opposite cerebellar hemisphere. It is supplied by the anterior inferior cerebellar artery (AICA) and branches from the basilar artery. It conveys information from the cerebrum and the pons to the cerebellum.

The pontocerebellar fibers are the second-order neuron fibers of the corticopontocerebellar tracts that cross to the other side of the pons and run within the middle cerebellar peduncles, from the pons to the contralateral cerebellum. They arise from the pontine nuclei as the second part of the corticopontocerebellar tract, and decussate (cross-over) in the pons before passing through the middle cerebellar peduncles to reach and terminate in the contralateral posterior lobe of the cerebellum (neocerebellum). It is part of a pathway involved in the coordination of voluntary movements.
The central tegmental tract is a tract that carries ascending and descending fibers, situated in the midbrain tegmentum, and the pontine tegmentum. The tract is situated in the central portion of the reticular formation.
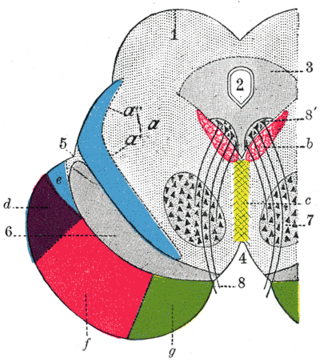
Corticopontine fibers are projections from layer V of the cerebral cortex to the pontine nuclei of the ventral pons. They represent the first link in a cortico-cerebello-cortical pathway mediating neocerebellar control of the motor cortex. The pathway is especially important for voluntary movements.
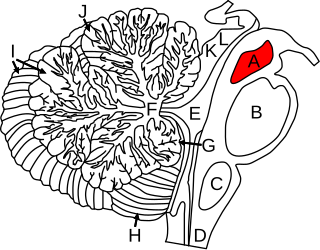
The anatomy of the cerebellum can be viewed at three levels. At the level of gross anatomy, the cerebellum consists of a tightly folded and crumpled layer of cortex, with white matter underneath, several deep nuclei embedded in the white matter, and a fluid-filled ventricle in the middle. At the intermediate level, the cerebellum and its auxiliary structures can be broken down into several hundred or thousand independently functioning modules or compartments known as microzones. At the microscopic level, each module consists of the same small set of neuronal elements, laid out with a highly stereotyped geometry.