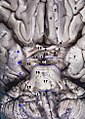
In neuroanatomy, the optic nerve, also known as the second cranial nerve, cranial nerve II, or simply CN II, is a paired cranial nerve that transmits visual information from the retina to the brain. In humans, the optic nerve is derived from optic stalks during the seventh week of development and is composed of retinal ganglion cell axons and glial cells; it extends from the optic disc to the optic chiasma and continues as the optic tract to the lateral geniculate nucleus, pretectal nuclei, and superior colliculus.

The visual system is the physiological basis of visual perception. The system detects, transduces and interprets information concerning light within the visible range to construct an image and build a mental model of the surrounding environment. The visual system is associated with the eye and functionally divided into the optical system and the neural system.

In neuroanatomy, the lateral geniculate nucleus is a structure in the thalamus and a key component of the mammalian visual pathway. It is a small, ovoid, ventral projection of the thalamus where the thalamus connects with the optic nerve. There are two LGNs, one on the left and another on the right side of the thalamus. In humans, both LGNs have six layers of neurons alternating with optic fibers.

The pupillary light reflex (PLR) or photopupillary reflex is a reflex that controls the diameter of the pupil, in response to the intensity (luminance) of light that falls on the retinal ganglion cells of the retina in the back of the eye, thereby assisting in adaptation of vision to various levels of lightness/darkness. A greater intensity of light causes the pupil to constrict, whereas a lower intensity of light causes the pupil to dilate. Thus, the pupillary light reflex regulates the intensity of light entering the eye. Light shone into one eye will cause both pupils to constrict.

A retinal ganglion cell (RGC) is a type of neuron located near the inner surface of the retina of the eye. It receives visual information from photoreceptors via two intermediate neuron types: bipolar cells and retina amacrine cells. Retina amacrine cells, particularly narrow field cells, are important for creating functional subunits within the ganglion cell layer and making it so that ganglion cells can observe a small dot moving a small distance. Retinal ganglion cells collectively transmit image-forming and non-image forming visual information from the retina in the form of action potential to several regions in the thalamus, hypothalamus, and mesencephalon, or midbrain.
In neuroanatomy, the optic tract is a part of the visual system in the brain. It is a continuation of the optic nerve that relays information from the optic chiasm to the ipsilateral lateral geniculate nucleus (LGN), pretectal nuclei, and superior colliculus.

The precentral gyrus is a prominent gyrus on the surface of the posterior frontal lobe of the brain. It is the site of the primary motor cortex that in humans is cytoarchitecturally defined as Brodmann area 4.
Axon guidance is a subfield of neural development concerning the process by which neurons send out axons to reach their correct targets. Axons often follow very precise paths in the nervous system, and how they manage to find their way so accurately is an area of ongoing research.

The abducens nucleus is the originating nucleus from which the abducens nerve (VI) emerges—a cranial nerve nucleus. This nucleus is located beneath the fourth ventricle in the caudal portion of the pons near the midline, medial to the sulcus limitans.

The facial motor nucleus is a collection of neurons in the brainstem that belong to the facial nerve. These lower motor neurons innervate the muscles of facial expression and the stapedius.

The nucleus of the trochlear nerve is a motor nucleus in the medial midbrain giving rise to the trochlear nerve.

Decussation is used in biological contexts to describe a crossing. In Latin anatomical terms, the form decussatio is used, e.g. decussatio pyramidum.
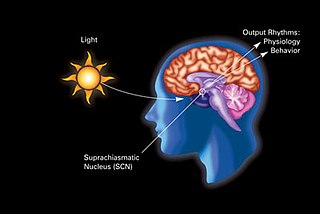
In neuroanatomy, the retinohypothalamic tract (RHT) is a photic neural input pathway involved in the circadian rhythms of mammals. The origin of the retinohypothalamic tract is the intrinsically photosensitive retinal ganglion cells (ipRGC), which contain the photopigment melanopsin. The axons of the ipRGCs belonging to the retinohypothalamic tract project directly, monosynaptically, to the suprachiasmatic nuclei (SCN) via the optic nerve and the optic chiasm. The suprachiasmatic nuclei receive and interpret information on environmental light, dark and day length, important in the entrainment of the "body clock". They can coordinate peripheral "clocks" and direct the pineal gland to secrete the hormone melatonin.

Chiasmal syndrome is the set of signs and symptoms that are associated with lesions of the optic chiasm, manifesting as various impairments of the affected's visual field according to the location of the lesion along the optic nerve. Pituitary adenomas are the most common cause; however, chiasmal syndrome may be caused by cancer, or associated with other medical conditions such as multiple sclerosis and neurofibromatosis.

This article describes anatomical terminology that is used to describe the central and peripheral nervous systems - including the brain, brainstem, spinal cord, and nerves.

The contralateral organization of the forebrain is the property that the hemispheres of the cerebrum and the thalamus represent mainly the contralateral side of the body. Consequently, the left side of the forebrain mostly represents the right side of the body, and the right side of the brain primarily represents the left side of the body. The contralateral organization involves both executive and sensory functions. The contralateral organization is only present in vertebrates.
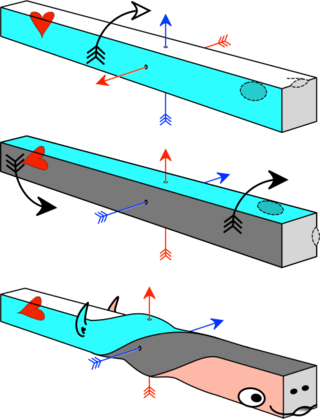
The axial twist theory is a scientific theory put forward to explain a range of unusual aspects of the body plan of vertebrates. It proposes that the rostral part of the head is "turned around" regarding the rest of the body. This end-part consists of the face as well as part of the brain. According to the theory, the vertebrate body has a left-handed chirality.
Carol Ann Mason is a Professor of Pathology and Cell Biology at Columbia University in the Mortimer B. Zuckerman Mind Brain Behavior Institute. She studies axon guidance in visual pathways in an effort to restore vision to the blind. Her research focuses on the retinal ganglion cell. She was elected a member of the National Academy of Sciences in 2018.

In anatomy a chiasm is the spot where two structures cross, forming an X-shape. Examples of chiasms are:

The visual pathway consists of structures that carry visual information from the retina to the brain. Lesions in that pathway cause a variety of visual field defects. In the visual system of human eye, the visual information processed by retinal photoreceptor cells travel in the following way:
Retina→Optic nerve→Optic chiasma →Optic tract→Lateral geniculate body→Optic radiation→Primary visual cortex