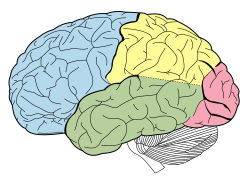
The parietal lobe is one of the four major lobes of the cerebral cortex in the brain of mammals. The parietal lobe is positioned above the temporal lobe and behind the frontal lobe and central sulcus.

In neuroanatomy, the precuneus is the portion of the superior parietal lobule on the medial surface of each brain hemisphere. It is located in front of the cuneus. The precuneus is bounded in front by the marginal branch of the cingulate sulcus, at the rear by the parieto-occipital sulcus, and underneath by the subparietal sulcus. It is involved with episodic memory, visuospatial processing, reflections upon self, and aspects of consciousness.

The motor cortex is the region of the cerebral cortex involved in the planning, control, and execution of voluntary movements. The motor cortex is an area of the frontal lobe located in the posterior precentral gyrus immediately anterior to the central sulcus.
A gamma wave or gamma rhythm is a pattern of neural oscillation in humans with a frequency between 25 and 140 Hz, the 40 Hz point being of particular interest. Gamma rhythms are correlated with large-scale brain network activity and cognitive phenomena such as working memory, attention, and perceptual grouping, and can be increased in amplitude via meditation or neurostimulation. Altered gamma activity has been observed in many mood and cognitive disorders such as Alzheimer's disease, epilepsy, and schizophrenia.
Visual search is a type of perceptual task requiring attention that typically involves an active scan of the visual environment for a particular object or feature among other objects or features. Visual search can take place with or without eye movements. The ability to consciously locate an object or target amongst a complex array of stimuli has been extensively studied over the past 40 years. Practical examples of using visual search can be seen in everyday life, such as when one is picking out a product on a supermarket shelf, when animals are searching for food among piles of leaves, when trying to find a friend in a large crowd of people, or simply when playing visual search games such as Where's Wally?

The premotor cortex is an area of the motor cortex lying within the frontal lobe of the brain just anterior to the primary motor cortex. It occupies part of Brodmann's area 6. It has been studied mainly in primates, including monkeys and humans. The functions of the premotor cortex are diverse and not fully understood. It projects directly to the spinal cord and therefore may play a role in the direct control of behavior, with a relative emphasis on the trunk muscles of the body. It may also play a role in planning movement, in the spatial guidance of movement, in the sensory guidance of movement, in understanding the actions of others, and in using abstract rules to perform specific tasks. Different subregions of the premotor cortex have different properties and presumably emphasize different functions. Nerve signals generated in the premotor cortex cause much more complex patterns of movement than the discrete patterns generated in the primary motor cortex.

The intraparietal sulcus (IPS) is located on the lateral surface of the parietal lobe, and consists of an oblique and a horizontal portion. The IPS contains a series of functionally distinct subregions that have been intensively investigated using both single cell neurophysiology in primates and human functional neuroimaging. Its principal functions are related to perceptual-motor coordination and visual attention, which allows for visually-guided pointing, grasping, and object manipulation that can produce a desired effect.

The supplementary motor area (SMA) is a part of the motor cortex of primates that contributes to the control of movement. It is located on the midline surface of the hemisphere just in front of the primary motor cortex leg representation. In monkeys the SMA contains a rough map of the body. In humans the body map is not apparent. Neurons in the SMA project directly to the spinal cord and may play a role in the direct control of movement. Possible functions attributed to the SMA include the postural stabilization of the body, the coordination of both sides of the body such as during bimanual action, the control of movements that are internally generated rather than triggered by sensory events, and the control of sequences of movements. All of these proposed functions remain hypotheses. The precise role or roles of the SMA is not yet known.
Eye–hand coordination is the coordinated motor control of eye movement with hand movement and the processing of visual input to guide reaching and grasping along with the use of proprioception of the hands to guide the eyes, a modality of multisensory integration. Eye–hand coordination has been studied in activities as diverse as the movement of solid objects such as wooden blocks, archery, sporting performance, music reading, computer gaming, copy-typing, and even tea-making. It is part of the mechanisms of performing everyday tasks; in its absence, most people would not be able to carry out even the simplest of actions such as picking up a book from a table.
The lateral intraparietal cortex is found in the intraparietal sulcus of the brain. This area is most likely involved in eye movement, as electrical stimulation evokes saccades of the eyes. It is also thought to contribute to working memory associated with guiding eye movement, examined using a delayed saccade task described below:
- A subject focuses on a fixation point at the center of a computer screen.
- A target is presented at a peripheral location on the screen.
- The target is removed and followed by a variable-length delay period.
- The initial focus point in the middle of the screen is removed.
- The subject's task is to make a saccade to the location of the target.
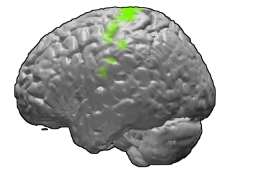
The primary motor cortex is a brain region that in humans is located in the dorsal portion of the frontal lobe. It is the primary region of the motor system and works in association with other motor areas including premotor cortex, the supplementary motor area, posterior parietal cortex, and several subcortical brain regions, to plan and execute voluntary movements. Primary motor cortex is defined anatomically as the region of cortex that contains large neurons known as Betz cells, which, along with other cortical neurons, send long axons down the spinal cord to synapse onto the interneuron circuitry of the spinal cord and also directly onto the alpha motor neurons in the spinal cord which connect to the muscles.
Visual object recognition refers to the ability to identify the objects in view based on visual input. One important signature of visual object recognition is "object invariance", or the ability to identify objects across changes in the detailed context in which objects are viewed, including changes in illumination, object pose, and background context.
Michael Steven Anthony Graziano is an American scientist and novelist who is currently a professor of Psychology and Neuroscience at Princeton University. His scientific research focuses on the brain basis of awareness. He has proposed the "attention schema" theory, an explanation of how, and for what adaptive advantage, brains attribute the property of awareness to themselves. His previous work focused on how the cerebral cortex monitors the space around the body and controls movement within that space. Notably he has suggested that the classical map of the body in motor cortex, the homunculus, is not correct and is better described as a map of complex actions that make up the behavioral repertoire. His publications on this topic have had a widespread impact among neuroscientists but have also generated controversy. His novels rely partly on his background in psychology and are known for surrealism or magic realism. Graziano also composes music including symphonies and string quartets.
Richard Alan Andersen is an American neuroscientist. He is the James G. Boswell Professor of Neuroscience at the California Institute of Technology in Pasadena, California. His research focuses on visual physiology with an emphasis on translational research to humans in the field of neuroprosthetics, brain-computer interfaces, and cortical repair.
Transsaccadic memory is the neural process that allows humans to perceive their surroundings as a seamless, unified image despite rapid changes in fixation points. Transsaccadic memory is a relatively new topic of interest in the field of psychology. Conflicting views and theories have spurred several types of experiments intended to explain transsaccadic memory and the neural mechanisms involved.
Form perception is the recognition of visual elements of objects, specifically those to do with shapes, patterns and previously identified important characteristics. An object is perceived by the retina as a two-dimensional image, but the image can vary for the same object in terms of the context with which it is viewed, the apparent size of the object, the angle from which it is viewed, how illuminated it is, as well as where it resides in the field of vision. Despite the fact that each instance of observing an object leads to a unique retinal response pattern, the visual processing in the brain is capable of recognizing these experiences as analogous, allowing invariant object recognition. Visual processing occurs in a hierarchy with the lowest levels recognizing lines and contours, and slightly higher levels performing tasks such as completing boundaries and recognizing contour combinations. The highest levels integrate the perceived information to recognize an entire object. Essentially object recognition is the ability to assign labels to objects in order to categorize and identify them, thus distinguishing one object from another. During visual processing information is not created, but rather reformatted in a way that draws out the most detailed information of the stimulus.
Gain field encoding is a hypothesis about the internal storage and processing of limb motion in the brain. In the motor areas of the brain, there are neurons which collectively have the ability to store information regarding both limb positioning and velocity in relation to both the body (intrinsic) and the individual's external environment (extrinsic). The input from these neurons is taken multiplicatively, forming what is referred to as a gain field. The gain field works as a collection of internal models off of which the body can base its movements. The process of encoding and recalling these models is the basis of muscle memory.

The ventrolateral prefrontal cortex (VLPFC) is a section of the prefrontal cortex located on the inferior frontal gyrus, bounded superiorly by the inferior frontal sulcus and inferiorly by the lateral sulcus. It is attributed to the anatomical structures of Brodmann's area (BA) 47, 45 and 44.
Social cognitive neuroscience is the scientific study of the biological processes underpinning social cognition. Specifically, it uses the tools of neuroscience to study "the mental mechanisms that create, frame, regulate, and respond to our experience of the social world". Social cognitive neuroscience uses the epistemological foundations of cognitive neuroscience, and is closely related to social neuroscience. Social cognitive neuroscience employs human neuroimaging, typically using functional magnetic resonance imaging (fMRI). Human brain stimulation techniques such as transcranial magnetic stimulation and transcranial direct-current stimulation are also used. In nonhuman animals, direct electrophysiological recordings and electrical stimulation of single cells and neuronal populations are utilized for investigating lower-level social cognitive processes.
Michael E. Goldberg, also known as Mickey Goldberg, is an American neuroscientist and David Mahoney Professor at Columbia University. He is known for his work on the mechanisms of the mammalian eye in relation to brain activity. He served as president of the Society for Neuroscience from 2009 to 2010.