In neuroscience and psychology, the term language center refers collectively to the areas of the brain which serve a particular function for speech processing and production. Language is a core system that gives humans the capacity to solve difficult problems and provides them with a unique type of social interaction. Language allows individuals to attribute symbols to specific concepts, and utilize them through sentences and phrases that follow proper grammatical rules. Finally, speech is the mechanism by which language is orally expressed.

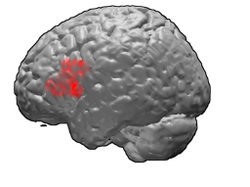
Broca's area, or the Broca area, is a region in the frontal lobe of the dominant hemisphere, usually the left, of the brain with functions linked to speech production.

The cingulate cortex is a part of the brain situated in the medial aspect of the cerebral cortex. The cingulate cortex includes the entire cingulate gyrus, which lies immediately above the corpus callosum, and the continuation of this in the cingulate sulcus. The cingulate cortex is usually considered part of the limbic lobe.

A Brodmann area is a region of the cerebral cortex, in the human or other primate brain, defined by its cytoarchitecture, or histological structure and organization of cells. The concept was first introduced by the German anatomist Korbinian Brodmann in the early 20th century. Brodmann mapped the human brain based on the varied cellular structure across the cortex and identified 52 distinct regions, which he numbered 1 to 52. These regions, or Brodmann areas, correspond with diverse functions including sensation, motor control, and cognition.

In neuroanatomy, the precuneus is the portion of the superior parietal lobule on the medial surface of each brain hemisphere. It is located in front of the cuneus. The precuneus is bounded in front by the marginal branch of the cingulate sulcus, at the rear by the parieto-occipital sulcus, and underneath by the subparietal sulcus. It is involved with episodic memory, visuospatial processing, reflections upon self, and aspects of consciousness.

The temporal lobe is one of the four major lobes of the cerebral cortex in the brain of mammals. The temporal lobe is located beneath the lateral fissure on both cerebral hemispheres of the mammalian brain.

Brodmann area 9, or BA9, refers to a cytoarchitecturally defined portion of the frontal cortex in the brain of humans and other primates. It contributes to the dorsolateral and medial prefrontal cortex.

Brodmann area 10 is the anterior-most portion of the prefrontal cortex in the human brain. BA10 was originally defined broadly in terms of its cytoarchitectonic traits as they were observed in the brains of cadavers, but because modern functional imaging cannot precisely identify these boundaries, the terms anterior prefrontal cortex, rostral prefrontal cortex and frontopolar prefrontal cortex are used to refer to the area in the most anterior part of the frontal cortex that approximately covers BA10—simply to emphasize the fact that BA10 does not include all parts of the prefrontal cortex.

Brodmann area 44, or BA44, is part of the frontal cortex in the human brain. Situated just anterior to premotor cortex (BA6) and on the lateral surface, inferior to BA9.
Brodmann area 46, or BA46, is part of the frontal cortex in the human brain. It is between BA10 and BA45.

Brodmann area 47, or BA47, is part of the frontal cortex in the human brain. It curves from the lateral surface of the frontal lobe into the ventral (orbital) frontal cortex. It is below areas BA10 and BA45, and beside BA11. This cytoarchitectonic region most closely corresponds to the gyral region the orbital part of inferior frontal gyrus, although these regions are not equivalent. Pars orbitalis is not based on cytoarchitectonic distinctions, and rather is defined according to gross anatomical landmarks. Despite a clear distinction, these two terms are often used liberally in peer-reviewed research journals.

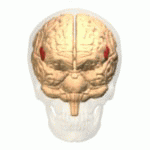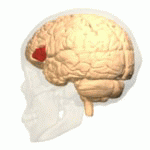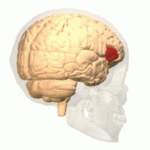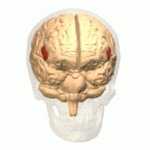
Wernicke's area, also called Wernicke's speech area, is one of the two parts of the cerebral cortex that are linked to speech, the other being Broca's area. It is involved in the comprehension of written and spoken language, in contrast to Broca's area, which is primarily involved in the production of language. It is traditionally thought to reside in Brodmann area 22, which is located in the superior temporal gyrus in the dominant cerebral hemisphere, which is the left hemisphere in about 95% of right-handed individuals and 70% of left-handed individuals.

Brodmann area 11 is one of Brodmann's cytologically defined regions of the brain. It is in the orbitofrontal cortex which is above the eye sockets (orbitae). It is involved in decision making, processing rewards, and encoding new information into long-term memory.

The inferior frontal gyrus (IFG),, is the lowest positioned gyrus of the frontal gyri, of the frontal lobe, and is part of the prefrontal cortex.

In neuroanatomy, the arcuate fasciculus is a bundle of axons that generally connects the Broca's area and the Wernicke's area in the brain. It is an association fiber tract connecting caudal temporal cortex and inferior frontal lobe.
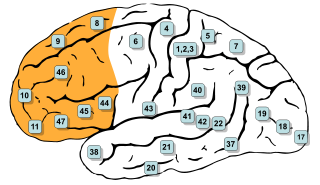
In mammalian brain anatomy, the prefrontal cortex (PFC) covers the front part of the frontal lobe of the cerebral cortex. It is the association cortex in the frontal lobe. The PFC contains the Brodmann areas BA8, BA9, BA10, BA11, BA12, BA13, BA14, BA24, BA25, BA32, BA44, BA45, BA46, and BA47.

Brodmann area 22 is a Brodmann's area that is cytoarchitecturally located in the posterior superior temporal gyrus of the brain. In the left cerebral hemisphere, it is one portion of Wernicke's area. The left hemisphere BA22 helps with generation and understanding of individual words. On the right side of the brain, BA22 helps to discriminate pitch and sound intensity, both of which are necessary to perceive melody and prosody. Wernicke's area is active in processing language and consists of the left Brodmann area 22 and Brodmann area 40, the supramarginal gyrus.

The lobes of the brain are the major identifiable zones of the human cerebral cortex, and they comprise the surface of each hemisphere of the cerebrum. The two hemispheres are roughly symmetrical in structure, and are connected by the corpus callosum. They traditionally have been divided into four lobes, but are today considered as having six lobes each. The lobes are large areas that are anatomically distinguishable, and are also functionally distinct to some degree. Each lobe of the brain has numerous ridges, or gyri, and furrows, the sulci that constitute further subzones of the cortex. The expression "lobes of the brain" usually refers only to those of the cerebrum, not to the distinct areas of the cerebellum.

The superior longitudinal fasciculus (SLF) is an association tract in the brain that is composed of three separate components. It is present in both hemispheres and can be found lateral to the centrum semiovale and connects the frontal, occipital, parietal, and temporal lobes. This bundle of tracts (fasciculus) passes from the frontal lobe through the operculum to the posterior end of the lateral sulcus where they either radiate to and synapse on neurons in the occipital lobe, or turn downward and forward around the putamen and then radiate to and synapse on neurons in anterior portions of the temporal lobe.
The orbital part of inferior frontal gyrus also known as the pars orbitalis is the orbital part of the inferior frontal gyrus.